#Eutherapsida
Text
Thrinaxodon

By Scott Reid
Etymology: Trident tooth
First Described By: Seeley, 1894
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota, Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Synapsida, Eupelycosauria, Metopophora, Haptodontiformes, Sphenacomorpha, Sphenacodontia, Pantherapsida, Sphenacodontoidea, Therapsida, Eutherapsida, Neotherapsida, Theriodontia, Eutheriodontia, Cynodontia, Epicynodontia, Thrinaxodontidae
Status: Extinct
Time and Place: Thrinaxodon lived 251 to 247 million years ago, in the Olenekian of the Early Triassic.

Thrinaxodon is known from South Africa and Antarctica.

Physical Description: If you thought Olivierosuchus was a good dog, Thrinaxodon is an even gooder dog. Like most synapsids around this area of the tree, Thrinaxodon would have been a small (~50 cm) badgery thing. Its head had whiskers, but probably didn’t have external ear pinnae (only holes in the back of the skull). The skull had a secondary palate that separated the nasal passages from the mouth, so it could continuously breathe while eating. Its teeth were divided into distinct incisors, canines, and molars, like modern mammals. The teeth were replaced continuously throuhhout its life, unlike most mammals (which get one tooth replacement, and that’s it). The inner ear still didn’t have the distinctive malleus and incus bones of mammalian middle ears, though. It held its limbs off the ground in a posture transitional between earlier semi-sprawling synapsids and straight-standing mammals. The forelimbs were not super-robust, but had features in common with modern digging mammals and reptiles. Each hand and foot had five digits each. Thrinaxodon probably had fur, although no direct fossil fur has been found yet.
Diet: Thrinaxodon’s diet likely consisted of small reptiles and invertebrates.
Behavior: As with the previously-featured Olivierosuchus, Thrinaxodon was able to burrow. We’ve found burrow casts with Thrinaxodon inside. Numerous Thrinaxodon specimens have been found curled up in little balls inside the burrows (it’s adorable), which suggests that it may have hibernated in these burrows during the heat of the summer. This may have helped cynodonts (as well as therocephalians) survive the Permian-Triassic extinction. In fact, one fossil burrow has both a curled-up Thrinaxodon and the temnospondyl Broomistega inside. As the Thrinaxodon was hibernating, an injured Broomistega crawled into the burrow to shelter itself and heal. It’s okay to cry at this, I’m doing so.
Thrinaxodon was a caring parent, looking out for clutches of its young until they were at least half full size. It would have laid soft-shelled eggs, like modern monotremes. Thrinaxodon is around the area of the synapsid tree where milk probably evolved into a thing that the babies drank for nutrients. Previously, “proto-milk” existed in the form of egg-moistening sweat. This said, the babies wouldn’t have suckled - instead, the milk would have been… sweated out of the mother’s skin, and they’d lick it off.
Ecosystem: Thrinaxodon fossils have been found in what were once arid floodplains in South Africa and Antarctica. At the time these were connected by land, so many animals lived in both places at the same time. Synapsids were prevalent in the region, such as Lystrosaurus (because of course), the cynodont Galesaurus, and the therocephalians Moschorhinus, Olivierosuchus, Ericiolacerta, Scaloposaurus, and Tetracynodon. Other animals that lived in South Africa include the parareptiles Procolophon and Owenetta, the early archosauromorphs Prolacerta and Proterosuchus, and temnospondyls such as Lydekkerina, Broomistega and Micropholis. A few species are known exclusively from Thrinaxodon’s Antarctic range, like the archosauriform Antarctanax and the dicynodont Kombuisia.
Other: Histology reveals that Thrinaxodon grew very fast, suggesting that it was warm-blooded. When young, Thrinaxodon grew quickly and replaced their teeth a lot. Growth rates slowed down considerably after a certain age, which may have been the point of sexual maturity.
~ By Henry Thomas
Sources under the cut
Abdala, F., Jasinoski, S.C., Fernandez, V. 2013. Ontogeny of the Early Triassic cynodont Thrinaxodon liorhinus (Therapsida): dental morphology and replacement. Journal of Vertebrate Paleontology 33(6): 1408-1431.
Botha, J., Chinsamy, A. 2005. Growth patterns of Thrinaxodon liorhinus, a non-mammalian cynodont from the lower Triassic of South Africa. Palaeontology 48(2): 385-394.
Brink, A.S. 1954. Note on a very tiny specimen of Thrinaxodon liorhinus. Palaeontologia Africana 3: 73-76.
Brink, A.S. 1959. Note on a new skeleton of Thrinaxodon liorhinus. Palaeontologia Africana 6: 15-22.
Damiani, R., Modesto, S., Yates, A., Neveling, J. 2003. Earliest evidence of cynodont burrowing. Proceedings of the Royal Society of London B 270: 1747-1751.
Estes, R. 1961. Cranial anatomy of the cynodont reptile Thrinaxodon liorhinus. Bulletin, Museum of Comparative Zoology, Harvard University 125: 165-180.
Fernandez, V., Abdala, F., Carlson, K.J., Cook, D.C., Rubidge, B.S., Yates, A., Tafforeau, P. 2013. Synchrotron Reveals Early Triassic Odd Couple: Injured Amphibian and Aestivating Therapsid Share Burrow. PLoS ONE 8(6): e64978.
Iqbal, S. 2015. The functional morphology and internal structure of the forelimb of the Early Triassic non-mammaliaform cynodont Thrinaxodon liorhinus. Unpublished dissertation, Evolutionary Studies Institute and School of Geosciences, University of the Wiwatersrand.
Jasinoski, S.C., Abdala, F. 2017. Aggregations and parental care in the Early Triassic basal cynodonts Galesaurus planiceps and Thrinaxodon liorhinus. PeerJ 5: e2875.
#thrinaxodon#thrinaxodon liorhinus#cynodont#synapsid#triassic#triassic madness#triassic march madness#prehistoric life#paleontology
206 notes
·
View notes
Text
Mammuthus

South African Mammoth by Scott Reid
Etymology: Earth-Horn
First Described By: Brookes, 1828
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Synapsida, Eupelycosauria, Sphenacodontia, Sphenacodontoidea, Therapsida, Eutherapsida, Neotherapsida, Theriodontia, Eutheriodontia, Cynodontia, Epicynodontia, Eucynodontia, Probainognathia, Chiniquodontoidea, Prozostrodontia, Mammaliaformes, Mammalia, Theriiformes, Holotheria, Trechnotheria, Cladotheria, Zatheria, Tribosphenida, Theria, Eutheria, Placentalia, Atlantogenata, Afrotheria, Paenungulata, Tethytheria, Proboscidea, Elephantiformes, Elephantimorpha, Elephantida, Elephantoidea, Elephantidae, Elephantinae, Elephantini, Elephantina
Referred Species: M. africanavus (African Mammoth), M. columbi (Columbian Mammoth), M. creticus (Cretan Dwarf Mammoth), M. exilis (Pygmy Mammoth), M. lamarmorai (Sardinian Pygmy Mammoth), M. meridionalis (Southern Mammoth), M. primigenius (Woolly Mammoth), M. rumanus (Romanian Mammoth), M. subplanifrons (South African Mammoth), M. trogontherii (Steppe Mammoth)
Status: Extinct
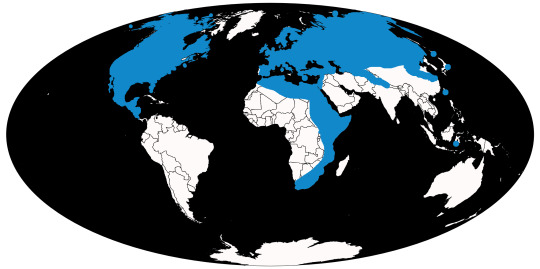
Time and Place: Mammoths have been around from 5 million years ago, until 3,700 years ago (give or take a few hundred years)


Mammoths are known from throughout North America and Afroeurasia
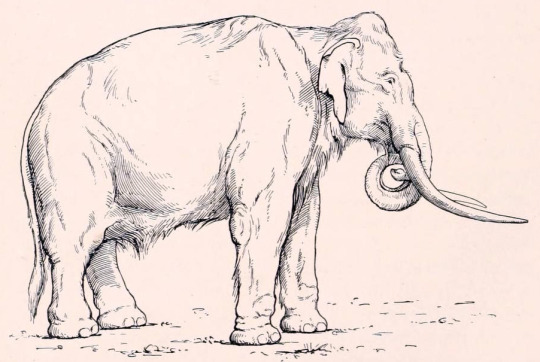
Physical Description: Mammoths were (mostly) large, elephanty creatures - with big bodies, long limbs, trunks, small tails, and tusks coming out of their mouths in both sexes. Most were about as large as Asian elephants, though some may have exceeded 12 tonnes. Still, there were dwarf species as well, which were quite small - some not even breaking 1000 kilograms. They grew their first tusks at the age of six months, which were then replaced at eighteen months by the permanent set. These would then grow at about 2.5 to 15.2 centimeters per year. Their teeth were distinctive - with rows of ridges that looked almost like the body of an arthropod (rather than the more flat surface seen in contemporary mastodons).

By FunkMonk, in the Public Domain
The mammoths had small ears, primarily due to the need to conserve body heat in most cases, unlike modern elephants which have larger ears with which to let off excess body heat. They had wide feet, splayed apart to help hold up their bodies. The males usually would grow larger than the females, based on the shape of the pelvis in fossils and frozen remains of these mammals. And, though a few species are very hairy - leading to the famed Woolly Mammoth - not all of them were, and many species of this genus were naked, with grey or more beige colored skin, as they lived in drier and warmer environments and needed to let off heat and sweat. Those that were hairy had thick layers of fur covering their entire bodies, even shorter tails to reduce frostbite, and thick layers of fat for keeping warm. A few species - notably, the Woolly Mammoth - had very brown, fluffy fur. Finally Mammoths, like modern elephants, were extremely intelligent animals, and had the brain and head size to match.

Woolly Mammoth by Mauricio Antón, CC BY 2.5
Diet: Mammoth diet varied from species to species and location to location of mammoths, though they were, overall, herbivores. Some mainly grazed on things like cacti leaves, trees, and shrubs; others, herbs, grasses, and shrubs; still more, forbs, rather than grass. Sometimes, babies that were no longer drinking milk would eat the poop of adults, like modern elephants, as they could not chew properly on plant material yet. This would also lead to feeding on fungi associated with poop, in addition to the plant material present.
Behavior: Mammoths were, in general, herding animals, much like modern elephants - in fact, it is theorized that they probably had social structures very similar to their living relatives. They had females living in herds, headed by a matriarch, that was the source of wisdom for the whole heard; meanwhile, the males would live alone or form loose groups of convenience when the situation arose. These herds would, oftentimes, migrate from region to region based on weather patterns and seasonal change. They could form herds of thousands of individuals of a variety of ages, with the adorable image of juveniles and young running to keep up with the strides of the much larger adults.

Woolly Mammoth by Flying Puffin, CC BY-SA 2.0
Mammoths, like modern elephants, mainly interacted with their environments with their trunks, which were essentially fifth limbs for these animals. They grabbed objects in their environments and talked to each other through the movement of their trunks. The trunks were also used for grabbing food in a lot of situations, pulling up coarse tundra grass and other food items and brought to the mouth, where it would then be chewed with the trough, specialized teeth. They used their tusks for defense from predators, though this wasn’t very helpful for the juveniles, who did not have extensive tusks. The mammoths also fought each other with their tusks, especially males, who would interlock the tusks and even break them while fighting for mates. They could also use the tusks to strip off bark and gather up food, even by digging in the dirt for food.
Ecosystem: Mammoths lived in a wide variety of habitats overall, though of course these differed from species to species. They lived across savanna and desert, steppe and tundra, grassland and scrubland, and even in sparse forest and higher elevations. As for predators, Mammoths were heavily at risk from predators such as lions, sabre-toothed cats, various types of wolves and other dogs, and of course, humans - which are probably the reason Mammoths are extinct today.

Cave Art from PalaeoAmerica, in the Public Domain, depicting two Columbian Mammoths (with a bison drawn over the right one)

Kapova Cave, in the Public Domain
Other: How did Mammoths go extinct? Apart from earlier species, the answer seems to be a combination of climate change and human activity. Multiple species - especially the Columbian and Woolly Mammoths - are known to have been hunted by humans, and cave drawings depict this activity for both of these species. This combination had a terrible effect on all species of mammoths - rising sea levels, with wildfires, and human hunting lead to the extinction of the Pygmy Mammoth, despite not being as large and specialized as those more iconic, giant species of mammoths elsewhere. Habitat shrinkage did not help with Mammoth populations; but human activity definitely contributed, as many populations of mammoth were heavily impacted by human activity. Prior to this, there was extensive genomic meltdown, as DNA diversity went into heavy decline for the past few thousand years. Isolated populations of mammoths - such as those on islands - showed the most extreme decline as the climate changed and humans became more common. The final death knell came from population fragmentation - with mammoths no longer having overlapping ranges, isolated populations were more susceptible to these rapid changes, and each subsequently went extinct due to climate change and human activity (with different causes being more or less important depending on the population in question).

Steppe Mammoth by Titus 322, in the Public Domain
Species Differences: There are 10 species of mammoth, each very distinct from the others, and each occupying unique environments and locations. Some are older than the others, as well.
The oldest species, M. subplanifrons, or the South African Mammoth, appeared about 5 million years ago - at the very beginning of the Pliocene. It spread from South Africa to the East African region, where it became most common in Ethiopia (funnily enough, the same time and location as human evolution, really). They were very similar to later mammoths, having spirally twisting tusks and large size - weighing up to 9 tonnes. They went extinct within the Early Pliocene and, given that they lived in Africa, were probably not very hairy as far as mammoths go; living mainly on savannas and other habitats that we see modern African Elephants on today.

African Mammoth by Apokryltaros, CC BY 2.5
The second oldest species, M. africanavus, the African Mammoths, lived from the Late Pliocene (about 3 million years ago) through the earliest part of the Pleistocene, about 1.65 million years ago. These two early species in Africa point to an African origin for the Mammoths. It has been found in Chad, Libya, Morocco, and Tunisia - showing the northern migration of Mammoths in their early evolution. They had very widely diverging tusks, much more so than other mammoths.
The next in the group, M. rumanus, the Romanian Mammoth, is one of our earliest examples of a Mammoth that spread to Europe - having been found in the late Pliocene of the UK and Romania. Being a very short lived and poorly known species, it is difficult to know much about it, though it does point to Mammoths migrating out of Africa through the Levant region, up to Eurasia from there. It’s possible that the Romanian Mammoth helps to show more about the dispersal of this genus through Eurasia as the Ice Age began in the Pleistocene.
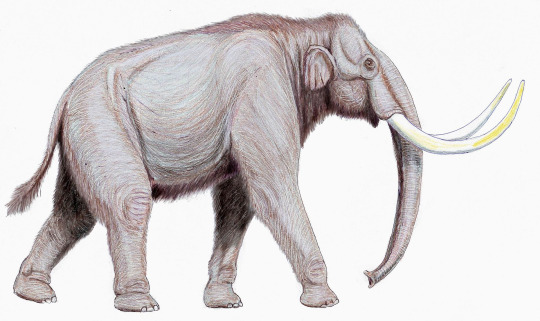
Southern Mammoth by Erwin S. Christman, in the Public DOmain
The Southern Mammoth, M. meridionalis, lived from the end of the Pliocene through the beginnings of the Pleistocene, and is known from Europe through Central Asia. It is known from many remains, which pinpoint that it may have been very closely related to the African Mammoth, though currently it is hypothesized that the South African Mammoth was the ancestor of all other forms. The Southern Mammoth grew to be about 4 meters tall and weighed 10 tonnes, making it one of the largest of the group. It had very robust, twisted tusks like other mammoths. It lived, generally, during a more mild time of the Ice Age, feeding mainly on deciduous trees and living in grassy, open habitats with small groves here and there. It was a browser, feeding mainly on higher level foliage.
Steppe Mammoth by Dmitry Bogdanov, CC BY 3.0
All other Mammoths lived from the Pleistocene on, with some making it into the Holocene (and subsequently going extinct during the Pleistocene-Holocene Megafaunal Extinctions). The Steppe Mammoth is the next in our temporal sequence, M. trogontherii, known from Siberia in the middle of the Pleistocene - the first stage in the evolution of steppe and tundra mammoths during the glacial periods of the time. It probably had fur on most of its body - shorter than that of the Woolly Mammoth, but not by much. It had a short skull and small jaw compared to other mammoths, and the males had tusks with tips that could grow extra long and curved. They grew up to 4 meters tall and around 10 tonnes in weight. It is known mainly from fossilized teeth, with skeletons being rare, though some complete skeletons have been excavated in northern parts of the UK and siberia. It probably ate mainly shrub plants and tundra forbs.

Columbian Mammoth by Dmitry Bogdanov, CC BY-SA 4.0
The next species of Mammoth to evolve was the famed Columbian Mammoth, of the mid Pleistocene through Early Holocene of North America. Though it’s range was next to that of the Woolly Mammoth, they clearly divided North America between the two, with Woolly taking the northern half of the continent, and the Columbian mammoth taking the southern half. The Columbian Mammoth is known from a lot of fossils, across a variety of localities, and show the initial spread into North America by mammoths in an earlier in-between period of the Ice Age than the expansion of Woolly Mammoths later. It evolved from the Steppe Mammoth, which migrated across the Bering Strait into North America, down corridors available between glaciers through to the bulk of the area that would be the continental United States. The Columbian Mammoth grew to 4 meters in height and 10 tonnes in weight, making it larger than the Woolly Mammoth and the African Elephant. It also had fairly primitive teeth compared to other mammoths. It had tusks directed farther apart than those of other mammoths, and it had a longer tail. Since it lived in warmer habitats, it lacked a lot of the adaptations for the cold, and probably didn’t have much in the way of hair for keeping warm. Its tusks were excessively long, especially compared to modern elephants.
Many Columbian Mammoths are found in Elephant Graveyard fossil deposits, indicating areas where the bones of individuals would accumulate due to the movement of water. Additionally, many fossil remains are found in tar pit accumulations, in addition to sinkholes and other natural traps. Most fossils found in these sites are actually males, which were more likely to put themselves in danger than the females - lured to these holes by warm water and vegetation at the edges. These elephants would have needed to spend most of their day foraging, using their trunks to pull up grass, flowers, and other types of food. It’s possible that these mammoths could have reached even 80 years in age, growing for most of their lives.

Columbian Mammoth by Charles R. Knight, in the Public Domain
The Cretan Dwarf Mammoth, M. creticus, is one of three diminished species of Mammoths, known from about 700,000 years ago on the island of Crete in the Mediterranean. These mammoths were very small, reaching only 1 meter tall and weighing only 310 kilograms - making it the smallest of the mammoths. It is possible that this animal wasn’t a mammoth at all, but another type of proboscidean; studies are still out on that one. Skulls of this animal may have formed the basis for the Grecian myth of the Cyclops!
The Sardinian Pygmy Mammoth, M. lamarmorai, was another species of small mammoth which evolved about 450,000 years ago, and is known only from the island of Sardinia. It grew about 1.4 meters tall and weighed up to 550 kilograms. It is known from many fossils, but no complete skeletons, mainly along the west coast. It is actually very uncertain what sort of mammoth this species originated from; it seems possible that the Steppe Mammoth colonized Sardinia, and then experienced Island Dwarfism as it was isolated on the island during an in-between period.

Woolly Mammoth by Honymand, CC BY-SA 4.0
The second to last species of Mammoth to evolve is, indeed, the best known of them all - M. primigenius, the Woolly Mammoth. Woolly Mammoths lived around the Arctic circle, in northern Eurasia and North America. Evolving from the Steppe Mammoth sometime between 400,000 and 150,000 years ago, it lived all the way up until the recent past, about 4,000 years ago - leading to the common fun fact that the Pyramids were built while Mammoths were still alive! Given it’s late position in the fossil record and extreme numbers during the last ice age, we have so many fossils and frozen remains of this animal that we know much of its life history (and the Woolly Mammoth remains one of the most controversial examples of something we might actually bring back through de-extinction). Because of this abundance of remains, we know a lot about its life appearance and life history - in fact, it’s the prehistoric animal with the best known appearance.
Woolly Mammoths were - as the name suggests - woolly, covered in layers of fur all over their bodies which was brown in color, as well as very thick layers of fat, very small ears and short tails, and flaps of skin covering orifices to keep them warm. The mammoths had oil glands in their skin, which secreted into their hair and helped the hair in repelling water, and gave the hair a glossy sheen. There was long, coarse guard hair on the outer layer, covering curly under-wool underneath. Interestingly enough, the Woolly Mammoth also had some weird physical characteristics not related to the warmth, such as a very high, domed head, and a sloping back with a shoulder hump. These were more present in adults, and not visible in juveniles. Their tusks were asymmetrical and extremely varied, and weirdly enough, most of the length of the tusks was inside the mouth. They had four molars at a time, used for chewing very tough vegetation.

Woolly Mammoths by Kira Sokolvskaia, CC BY-SA 3.0
The Woolly Mammoth was the most specialized elephant to ever live, with extreme amounts of fat stored for warmth and when food was unavailable; their molars grew more quickly than in modern elephants; and their fur was so thick it was as though they wore mittens all over their bodies. They had differences in circadian rhythm clocks from living elephants in order to deal with extreme variation in daylight levels, and its proteins were less sensitive to heat. They had about 1.4 million differences in DNA compared to their closest living relatives, the Asian Elephants. They fed mainly on herbs, small flowering plants, shrubs, and mosses, and even fungus and poop from each other - not just the young, but the adults would do this too in times of food scarcity. They probably could reach up to 60 years of age, and they grew past adulthood - like living elephants.
The males also would go into musth - a period of extreme aggressiveness - during the breeding season, like living elephants today. They produced oil with glands that moved a smell associated with musth all over their fur, signaling to female mammoths they were ready to go, and to male mammoths to leave them alone. The breeding season was typically in the summer through the beginning of fall. The mammoths gave birth during the spring and summer, after a gestation period of nearly two years. While many reached older age and adulthood, bone disease was a very common cause of death for these mammals, as well as parasitic animals and infections after poorly healed injuries. Still, many were murdered by humans, and mammoth bits are used heavily in human society - for food, warmth, and art, with ivory from mammoths used in sculptures by early humans.

Pygmy Mammoth by Apokryltaros, CC BY 2.5
Finally, Pygmy Mammoths - M. exilis - were the last species to evolve, evolving approximately 60,000 years ago; still, it didn’t live as long as the Woolly Mammoth, dying out about 11,000 years ago, right at the beginning of the Holocene. The third case of insular dwarfism in the Mammoths, it grew to about 1.72 meters tall at the shoulders and 760 kilograms in weight. It evolved from the giant Columbian Mammoth, as well, making this all the more impressive. It is known from the Channel Islands along California, where Columbian Mammoths presumably reached via land bridges during some glacial period of the Ice Age. Possibly, they also swam there in search of food and escaping the large predators of the mainland such as Smilodon and the American Lion. It thrived across a variety of ecosystems, including plateaus, dunes, grasslands, forests, and even steppe-tundras.
~ By Meig Dickson
Sources under the Cut
https://en.wikipedia.org/wiki/Mammoth
https://en.wikipedia.org/wiki/African_mammoth
https://en.wikipedia.org/wiki/Columbian_mammoth
https://en.wikipedia.org/wiki/Mammuthus_creticus
https://en.wikipedia.org/wiki/Pygmy_mammoth
https://en.wikipedia.org/wiki/Mammuthus_lamarmorai
https://en.wikipedia.org/wiki/Mammuthus_meridionalis
https://en.wikipedia.org/wiki/Woolly_mammoth
https://en.wikipedia.org/wiki/Mammuthus_rumanus
https://en.wikipedia.org/wiki/Mammuthus_subplanifrons
https://en.wikipedia.org/wiki/Steppe_mammoth
#Mammuthus#Mammoth#Woolly Mammoth#Not a Dinosaur#Mammal#April Fool's#Prehistoric Life#Prehisotry#Paleontology#North America#Eurasia#Africa#Neogene#Quaternary#Herbivore#Ice Age#prehistory#biology#science#nature#factfile
861 notes
·
View notes
Video
Matschie's Tree Kangaroo (Dendrolagus matschiei) at Woodland Park Zoo (5) by Nick Michalski
#animal#Animalia#Chordata#Dendrolagus#Dendrolagus matschiei#deuterostome#Diprotodontia#Eugnathostomata#Eutherapsida#Huon Tree Kangaroo#macropod#Macropodidae#mammal#Mammalia#Mammaliamorpha#marsupial#Matschie's Tree Kangaroo#Sarcopterygii#tetrapod#Therapsida#Theriiformes#tree kangaro#tree-kangaroo#vertebrate#Woodland Park Zoo
0 notes
Text
Do You Love The Colour Of The Sky?
Eukaryote
Orthokaryotes
Neokaryotes
Scotokaryotes
Podiata
Unikonta
Obazoa
Opisthokonta
Holozoa
Filozoa
Animalia
Eumetazoa
ParaHoxozoa
Planulozoa
Bilateria
Nephrozoa
Deuterostomia
Chordata
Olfactores
Craniata
Vertebrata
Gnathostomata
Eugnathostomata
Teleostomi
Euteleostomi
Sarcopterygii
Rhipidistia
Tetrapodomorpha
Eotetrapodiformes
Elpistostegalia
Stegocephalia
Tetrapoda
Reptiliomorpha
Amniota
Synapsida
Eupelycosauria
Sphenacodontia
Sphenacodontoidea
Therapsida
Eutherapsida
Neotherapsida
Theriodontia
Eutheriodontia
Cynodontia
Epicynodontia
Eucynodontia
Probainognathia
Prozostrodontia
Mammaliaformes
Mammalia
Theriiformes
Holotheria
Trechnotheria
Symmetrodonta
Cladotheria
Zatheria
Tribosphenida
Theria
Eutheria
Placentalia
Boreoeutheria
Euarchontoglires
Euarchonta
Primatomorpha
Primates
Haplorhini
Simiiformes
Catarrhini
Hominoidea
Hominidae
Homininae
Hominini
Homo
sapiens
(something might be missing and I know some might be controversial)
0 notes
Text
Olivierosuchus parringtoni

By Ripley Cook
Etymology: Crocodile from Oliviershoek Pass
First Described By: Brink, 1965
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota, Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Synapsida, Eupelycosauria, Metopophora, Haptodontiformes, Sphenacomorpha, Sphenacodontia, Pantherapsida, Sphenacodontoidea, Therapsida, Eutherapsida, Neotherapsida, Theriodontia, Eutheriodontia, Therocephalia, Scylacosauria, Eutherocephalia, Akidnognathidae
Status: Extinct
Time and Place: Olivierosuchus lived 251 to 249 million years ago, in the Olenekian of the Early Triassic.
Olivierosuchus is only known from South Africa.

Physical Description: I feel like most of you would look at Olivierosuchus and think “that’s a good pupper”. Olivierosuchus had a fairly long snout that was slightly wider than deep. The snout has a slight upwards bend and has small pits that indicate the presence of whiskers. It had three types of teeth: average-sized incisors, a pair of large canine teeth, and small teeth in the back of the jaw. The body was low-slung and may have had semi-sprawling forelimbs. Its forelimbs were robust and were likely adapted for digging. Olivierosuchus had whiskers, and may have had a more extensive covering of fur.
Before you ask - Olivierosuchus was not venomous like has been suggested for its relative Euchambersia. Its teeth do not have grooves for spreading venom.
Diet: Olivierosuchus was carnivorous, and fed on smaller animals like parareptiles and the young of other synapsids.
Behavior: First - burrows can preserve in the fossil record, and it’s really interesting. Such as one large, wide burrow preserved in early Triassic rocks from South Africa. Interestingly, this burrow has the skeleton of a juvenile Lystrosaurus inside. The Lystrosaurus was too small to dig the burrow, but it’s the perfect size for something like Olivierosuchus. So an Olivierosuchus probably lived in the burrow, and brought back a dead Lystrosaurus juvenile as food. The back teeth of therocephalians suggests they may have been able to chew their food a bit before swallowing it. This is in contrast to gorgonopsians, which were forced to tear off swallowable chunks of food.
Ecosystem: Olivierosuchus lived alongside Lystrosaurus, because of course it did, everything alive at the beginning of the Triassic lived alongside Lystrosaurus. In fact the specific rock layer it was found in - which was deposited immediately after the Permian-Triassic Extinction - is called the Lystrosaurus Assemblage Zone. The environment was arid and not exactly lush, because of the extinction that just happened. The biostratigraphy of the Lystrosaurus Assemblage Zone is well-mapped enough that we know which therocephalians lived at exactly the same time as Olivierosuchus: those being Moscorhinus, “Tetracynodon” darti, Ericolacerta, Regisaurus, and Zorillodontops. The Lystrosaurus Assemblage Zone is also known for lots of Procolophon and Sauropareion, as well as cynodonts such as Thrinaxodon and Galesaurus. Early archosaurs are also known, such as Prolacerta and Proterosuchus, as well as temnospondyls like Broomistega and Lydekkerina.
Other: Olivierosuchus was originally named Olivieria by A.S. Brink in 1965. It was renamed Olivierosuchus in 2002 by Christian Kammerer and Christian Sidor, because Olivieria was already taken by a fly.
~ By Henry Thomas
Sources under the cut
Brink, A.S. 1965. A new Ictidosuchid (Scaloposauria) from the Lystrosaurus-Zone. Palaeontologia Africana 9: 129-138.
Kammerer, C.F., Sidor, C.A. 2002. Replacement names for the therapsid genera Criocephalus Broom 1928 and Olivieria Brink 1965. Palaeontologia Africana 38: 71-72.
Botha-Brink, J., Modesto, S.P. 2011. A new skeleton of the therocephalian synapsid Olivierosuchus parringtoni from the Lower Triassic South African Karoo Basin. Palaeontology 54(3): 591-606.
Modesto, S.P., Botha-Brink, J. 2010. A burrow cast with Lystrosaurus skeletal remains from the Lower Triassic of South Africa. Palaios 25(4): 274-281.
#olivierosuchus#akidnognathid#therocephalian#synapsid#triassic#triassic madness#triassic march madness#prehistoric life#paleontology
213 notes
·
View notes
Text
Chiniquodon
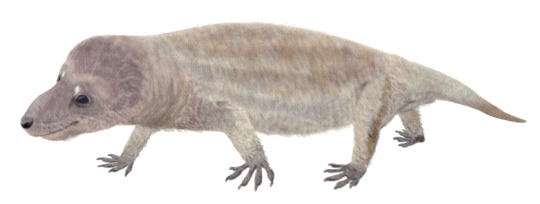
(Image: Chiniquodon, a medium-sized, rather generic-looking protomammal with a short tail, depicted here with a covering of short hair. Image by @Stolpergeist)
Etymology: Tooth from Chiniquá, Brazil
First described by: von Huene, 1936
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Synapsida, Eupelycosauria, Sphenacodontia, Sphenacodontoidea, Therapsida, Eutherapsida, Neotherapsida, Theriodontia, Eutheriodontia, Cynodontia, Probainognathia, Chiniquodontoidea, Chiniquodontidae
Referred Species: C. theotonicus, C. brasilensis, C. kalanoro, C. sanjuanensis
Status: Extinct (sadly :( )
Time and place: Chiniquodon lived from the Ladinian to the Carnian of South America and Africa (including Madagascar), about 242-227 million years ago.


Physical Description: As a cynodont, Chiniquodon was a close relative of mammals, but it’s aberrant compared to most fossil mammal relatives in one key respect: It’s known from more than just teeth! You heard that right, Chiniquodon is known from several reasonably complete skulls, as well as some body fossils. Thus, we can actually have some discussion of its appearance and anatomy beyond the teeth!
Chiniquodon was a medium to large (for the time) cynodont, with a skull length up to 33cm/13in in length. This skull had wide zygomatic arches (attachment points for jaw muscles!), and a solid secondary palate (as in modern mammals).
Its limbs and hips show that it was adapted to walk with its legs directly under the body, as in modern mammals. It had a fairly short tail. While we don’t have any direct evidence of its body covering, it’s likely that it had some hair, as has been inferred in other cynodonts. It got up to about the size of a medium dog.
…..But let’s be real, this is a mammal relative. We have to talk about teeth at some point.
Luckily for us, Chiniquodon has interesting teeth! Like, really interesting. Here’s a diagram of the skull of a typical chiniquodont:
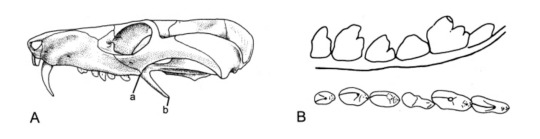
(Image: A diagram of the skull of a typical chiquodont. It’s got very large canines. Illustration B shows its cheek teeth from the side (top) and from the top (bottom). Image from Abdala and Giannini, 2002.)
What is the significance of these teeth? I guess you’ll have to wait until the “diet” section to find that out!
Diet: Oh look, we’re here. What were we on? Oh, right! The significance of these teeth. I’m sure many of you are chomping at the bit to find out what they were eating, and are about to point to those huge canines as proof that they were ravenous carnivorous beasts. And you may be on the cusp of an incisive argument, but I’d like to remind you that large canines do not a carnivore make, and modern musk deer have large canines themselves. To truly understand what an animal is eating, we need to look at the teeth that we know it used to process food. In this case, those are the cheek teeth.
The cheek teeth of Chiniquodon are a particular shape of tooth called a carnassial. That carn- is the same carn- as in carnivore or carnival or chile con carne, and it means meat. That’s because it’s a shape of cheek tooth that is especially good at cutting meat, because they have very large cusps that shear past each other when the animal bites down. Carnassials are found in many modern carnivorans (including dogs and cats!), but the carnassials of Chiniquodon evolved completely independently from those of dogs or cats – an amazing example of convergent evolution.
Behaviour: As a terrestrial carnivore, Chiniquodon must have hunted at some time. It likely hunted small vertebrates, such as other synapsids or young dinosaurs, though it likely wouldn’t have turned down scavenging from a carcass or eating bugs when it was younger. It almost certainly laid eggs, which it may have laid in a burrow. It is not clear whether any parental care was present.
Ecosystem: During the Triassic, all of the continents were combined into one, and this led to a very hot and dry climate over much of the Earth. Chiniquodon’s environment was no exception, and fossils of it are found in rocks interpreted to have been deposited by braided streams in an arid environment. With that said, some fossils of it are also found in rocks distributed during the Carnian Pluvial Event (aka That Time It Rained For Several Thousand Years; no relation to carnassials). These rocks contain the remains of animals like the early dinosaurs Eoraptor and Herrerasaurus, crocodile relatives like Gualosuchus, large dicynodonts like Ischigualastia, and other cynodonts like Probainognathus.
Other: Chiniquodon, in our prehistoric hindsight, is positioned right around one of the most significant transitions in mammalian evolution. It records the condition of cynodonts at a pivotal point, when it and its relatives were only just evolving furry body coverings and specialised jaw joints, and it’s a crucial fossil in untangling how its close close relatives miniaturised, going from medium-sized animals to the tiny shrew-like creatures that became the ancestors of all mammals. Today, with our prehistoric hindsight, Chiniquodon represents a sideshoot off of the mammal line during a pivotal transition time – but in its brightest day, when it was scurrying about the floodplains of South America and Africa, it was merely another animal fighting for existence in the world of the Triassic.
~ By Hayley Orlowski
Sources under the cut
Abdala, F.; Giannini, N. P. (2002). "Chiniquodontid cynodonts: systematic and morphometric considerations". Palaeontology. 45 (6): 1151–1170. doi:10.1111/1475-4983.00280.
Kammerer, C. F., Flynn, J. J., Ranivoharimanana, L., & Wyss, A. R. (2010). The first record of a probainognathian (Cynodontia: Chiniquodontidae) from the Triassic of Madagascar. Journal of Vertebrate Paleontology, 30(6), 1889–1894. doi:10.1080/02724634.2010.520784
Kemp, T. S. (2009). The endocranial cavity of a nonmammalian eucynodont, Chiniquodon theotenicus, and its implications for the origin of the mammalian brain. Journal of Vertebrate Paleontology, 29(4), 1188–1198. doi:10.1671/039.029.0430
Martinez, R. N., & Forster, C. A. (1996). The skull of Probelesodon sanjuanensis, sp. nov., from the Late Triassic Ischigualasto Formation of Argentina. Journal of Vertebrate Paleontology, 16(2), 285–291. doi:10.1080/02724634.1996.10011315
Mocke, H. B. (2016). The postcranium of the carnivorous cynodont Chiniquodon from the Middle Triassic of Namibia and the palaeo-environment of the Upper Omingonde Formation (Masters dissertation).
Oliveira, T. D., Schultz, C. L., & Soares, M. B. (2009). A partial skeleton of Chiniquodon (Cynodontia, Chiniquodontidae) from the Brazilian Middle Triassic. Revista Brasileira de Paleontologia, 12(2), 113-122.
#chiniquodon#chiniquodontian#cynodont#probainognathian#triassic#triassic madness#triassic march madness#prehistoric life#paleontology
257 notes
·
View notes
Text
Lystrosaurus

By Jack Wood / Darwin’s Door
Etymology: Shovel reptile
First Described By: Cope, 1870
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota, Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Synapsida, Eupelycosauria, Sphenacodontia, Sphenacodontoidea, Therapsida, Eutherapsida, Neotherapsida, Anomodontia, Chainosauria, Dicynodontia, Therochelonia, Bidentalia, Dicynodontoidea, Lystrosauridae
Referred Species: L. curvatus, L. declivis, L. georgi, L. hedini, L. maccaigi, L. murrayi (and several other probably invalid species)
Status: Extinct
Time and Place: Between 253 and 247 million years ago, from the Changhsingian of the Late Permian to the Olenekian of the Early Triassic

Lystrosaurus is mostly known from the Karoo Basin in South Africa, along with the Fremouw Formation in Antarctica, the Panchet Formation in India, and probably Australia in Gondwana, while at least two or more species from Laurasia are known in Russia, China, and Mongolia, giving Lystrosaurus a very broad distribution across much of Pangaea.

Physical Description: Lystrosaurus is pretty much the quintessential dicynodont, but when you get down to it, it’s actually a pretty weird one. The skull is unusually tall and squashed looking, even by dicynodont standards, with an incredibly short snout that drops down very steeply almost right in front of the eyes to the squared-off beak and pointy tusks at the bottom, hence the resemblance to a shovel (although the face of Lystrosaurus is actually surprisingly narrow from the front). In some specimens, the snout drops down almost vertically. The eyes are placed so far forward and up on the skull that they almost look like they’re bugging out of its head, and the nostrils are sat directly below them on the front of the snout. The back region of the skull behind the eyes takes up at least half of the length of the skull and houses massive jaw muscles, giving Lystrosaurus a very strong, snapping bite. Curiously, at least some Lystrosaurus may have had a thickened cornified pad between the eyes, possibly similar to the keratin of the beak, as well as rough bosses over the eyes. These were probably used for display, and may have been surprisingly colourful for what you’d expect from a stem-mammal.
The body of Lystrosaurus is more standard, with a long, tubular trunk, stubby tail and short, squat legs. Overall it was pretty dumpy looking (this is an actual description used in a scientific paper, no joke). This shape has been compared to that of hippos, often to suggest a similar lifestyle, but the strong, semi-upright limbs suggest a more terrestrial lifestyle. The forelimbs of Lystrosaurus are very well developed, and its hands and feet are broad with spread toes and large, flat claws, well suited for digging. It was somewhere around the size of the pig, give or take how big you think a pig is, but since Lystrosaurus sizes vary between small to over a metre you’re probably in the ballpark. The integument of Lystrosaurus, and indeed almost all stem-mammals, is poorly understood, and they have been speculated to be scaley, hairy, or just covered in naked skin. However, as-yet unpublished Lystrosaurus mummies from South Africa suggest that Lystrosaurus had hairless, scaleless skin studded with bumpy tubercles. But until we know more about these specimens, we can’t be certain how this would look. Lystrosaurus may also have been sexually dimorphic, with presumed males having more prominent bosses and robuster skulls. Lystrosaurus grew fast for dicynodonts, with rapid continuous growth as juveniles, slowing down as sub-adults with periods of little to no growth, and slowing considerably as they reached adulthood.
Diet: Lystrosaurus was a herbivore, grazing mostly on tough, low-lying drought-resistant plants like horsetails and seed ferns. However, it is also possible that it ate fungi and even bits of animal matter too while rooting around in the earth. Like other dicynodonts, Lystrosaurus chewed its food with a backwards motion of the jaw using its massive jaw muscles, making it an efficient grazer, and its short skull and jaws made it extra effective at snapping through tough stems.
Behavior: Lystrosaurus has often been described as amphibious, living like a hippopotamus, based on its supposedly high set eyes and downward facing jaws. However, there is actually little evidence to support this mode of life, and in reality the evidence is more in favour of a mostly terrestrial lifestyle, no more aquatic than any other dicynodont. Bonebeds of Lystrosaurus tells us that they were social animals that at least sometimes congregated together in similar age groups, at least as juveniles and subadults, possibly for protection during extreme temperature swings. For a long time Lystrosaurus was speculated to have been a burrower, and this was confirmed later by the discovery of not only Lystrosaurus burrows, but even burrows with the skeleton of its owner still inside. Burrowing was evidently a successful strategy for surviving the Great Dying, as many other Early Triassic survivors, like Thrinaxodon and Procolophon, were also burrowers. In fact, it’s even possible that Lystrosaurus acted as a sort of ecosystem engineer, modifying the environment and providing other species with refugiums in hard times that allowed them to pull through alongside it!
Ecosystem: In the earliest Triassic, Lystrosaurus was famously one of the most abundant vertebrates on the planet. It is regarded as a disaster taxon, a species that survives and proliferates in the ecological vacuum left after a mass extinction. Because the continents were joined together in Pangaea, much of the world’s fauna and flora was broadly similar. This is especially so in Gondwana, where much of the therapsid, reptile and amphibian fauna was shared. Here Lystrosaurus co-existed with various other therapsids, including two other surviving dicynodonts Myosaurus and Kombuisia, as well as the small cynodonts Thrinaxodon and Galesaurus, and various therocephalians such as Ericiolacerta, Tetracyndon, Ictidosuchus and the 1.5 metre long predatory Moschorhinus, one of the few predators that could prey upon Lystrosaurus in the Early Triassic. Archosauromorphs had also survived here, most notably the predatory proterosuchids, as well as the small Prolacerta and the larger, exclusively Antarctic Antarctax. Other reptiles includes the parareptile Procolophon, and the enigmatic diapsid Palacrodon, while the closely related (and relatively terrestrial as it turns out) temnospondyls Lydekkerina and Cryobatrachus cruised the waterways of South Africa and Antarctica, respectively. Similar fossils are known from India as well.
Prior to the P-Tr Extinction, in the time of L. maccaigi, South Africa was still relatively wet and lush with vegetated floodplains of trees and shrubs, including the famous Glossopterus seed ferns that L. maccaigi may have specialised in feeding on. However, this environment was already changing as the P-Tr Extinction took course, and was already low in biodiversity. L. maccaigi would have coexisted with other dicynodonts, including the lystrosaurid Kwazulusaurus, larger Daptocephalus, Oudenodon, Pelanomodon, and Dinanomodon, and little Thliptosaurus. Predatory therocephalians were around too, namely Promoschorhinus and Moschorhinus itself, which was another P-Tr survivor like Lystrosaurus. Most others however would be lost to the Great Dying, as the Karoo gradually dried out until it was transformed into a hot, arid plain with sprawling braided rivers, little vegetation and seasonally extreme temperatures that was prone to flash floods and cold snaps. To give a clear picture of how arid it was, it’s speculated that so many Lystrosaurus skeletons are found partially articulated and not broken apart like so many other Karoo fossils is because their carcasses were dessicated into mummies that held them together before they were eventually buried whenever it actually rained.
Lystrosaurus may have been pre-adapted to such conditions, however, as its downturned jaws would have been well-suited to grazing on low-growing drought-resistant plants, such as horsetails, as well as for Dicroidium seed ferns that would soon become the dominant Triassic flora. The Fremouw Formation in Antarctica records a more diverse range of seed ferns, cycads, horsetails and even fungi, all of which may have sustained Lystrosaurus. The climate at these high latitudes was milder, but also would mean they would have experienced months of darkness during the polar winters, even if it wasn’t cold.
Other: Lystrosaurus fossils were pivotal in proving Alfred Wegner’s theory of plate tectonics, as they had been found on various continents around the globe, along with other similarly aged fossils, indicating that the continents had once been much closer together and connected as one landmass. Their fossils are famously ludicrously abundant in the Karoo basin, making one it one of the best represented dicynodonts known, and probably of any stem-mammal. In fact, their abundance is so characteristic of the Early Triassic beds of the Karoo that they lend it its name, the Lystrosaurus Assemblage Zone. Lystrosaurus is sometimes regarded as one of the most successful tetrapods of all time, due to how widespread and populous it was. Case in point, 73% of known vertebrate fossils from the Early Triassic of South Africa are of Lystrosaurus. It’s a prime example of how a disaster taxon with the right pre-adaptations can flourish in the immediate aftermath of a mass extinction, and help to re-establish biotic communities during the recovery of global ecosystems. Not bad for a pudgy, dumpy little herbivore with a squashed up face.
Species Differences: To be frank, and I’m sure most synapsid workers won’t mind me saying this, the majority of stem-mammal taxonomy, including Lystrosaurus, is heavily biased towards their skulls. So much so that the majority of Lystrosaurus species were named based solely on slight variations between their skulls and so consequently were massively oversplit. Thankfully this is being rectified with more thorough, dedicated examinations of Lystrosaurus specimens, and so far the Gondwanan species of Lystrosaurus in the Karoo are the best studied. Four species from there are now recognised, and by and by they’re all still pretty samey looking, superficially. These four species have been recognised as sequentially spanning across the Late Permian and into the Early Triassic, and so can also be distinguished by when they each lived.
L. maccaigi is both the largest and the oldest species, and is the only one known exclusively from the Permian (although a single skull from Antarctica may or may not bring it into the earliest Triassic, assuming the rocks weren’t really Permian all along). It’s recognised for having remarkably well-developed bosses both in front and behind the eyes, giving it very distinctive looking “brows”, and the eyes themselves are characteristically large and placed high on the skull, facing somewhat more forwards and upwards than other species. The snout also drops down especially sharply (almost vertically in some), with a straight edge and ridge down the centre. As well as being the largest species, L. maccaigi was also the rarest, and both of these factors may have contributed to its extinction in the P-Tr.
The smaller L. curvatus is also known from the Permian, but unlike L. maccaigi it is known to have actually crossed the boundary into the earliest Triassic, however it died out shortly afterwards. L. curvatus is one of the least ornamented species of Lystrosaurus, with very reduced or no ridges and bosses on the face and over the eyes, although the largest individuals may have them, including small nasal bosses. L. curvatus also has some of the proportionally largest eyes of all Lystrosaurus, as well as a more gently curved snout than the others. Probably the cutest looking.
L. declivis and L. murrayi are exclusively found in the Triassic, and interestingly they are both consistently smaller than either Permian species. This is believed to be a response to the harsh conditions of the Permian Extinction and Early Triassic, as Lystrosaurus were growing faster and reaching reproductive maturity at smaller sizes and younger ages to combat shorter life expectancies. They are both fairly similar in appearance, each with short, slightly curved, somewhat angular snouts, although L. murrayi has a shorter snout only as long as the roof of the skull, whereas L. declivis has a snout that extends further down. L. declivis also has a pair of bosses in front of the eyes and a distinct ridge running between them, as well as a ridge running down the beak, all features missing from L. murrayi. Both species have a patch of grooves between the eyes on the roof of the skull, possibly supporting a sheet of keratin not seen in L. maccaigi or L. curvatus.
However, this apparent timeline does not mean there was a linear progression between these species. In fact, L. maccaigi and L. curvatus are the most derived species of Lystrosaurus, while L. declivis and L. murrayi are each more primitive than the last! Perhaps the more primitive species were able to survive by being less specialised and growing smaller. Also, this means that their ancestors had already split off during the Permian before L. maccaigi even evolved, so Gondwanan Lystrosaurus did not survive the P-Tr Extinction in just one species, but at least three! L. curvatus and L. murrayi are also both known from Triassic Antarctica, and L. murrayi is further known from India, implying quite a broad Gondwanan distribution—although given the three continents’ close proximity in Pangaea this may not be surprising.
The differences between the Laurasian species are less clear, and while various species have been named from China, it is possible that they belong to only a single species, L. hedini. L. hedini is another species known from the Permian to have crossed into the Early Triassic, and has also been found in Mongolia.
L. georgi from the Triassic of Russia is a bit of an enigma, unlike virtually every other Lystrosaurus species its skeleton is better known than its skull. This is great for understanding how the body of Lystrosaurus functioned but less so for its taxonomy. In any case, L. georgi is still pretty biogeographically distinct from other Lystrosaurus species, so it has merit. L. georgi was initially regarded as similar to the Gondwanan L. curvatus, but preliminary analyses of other Laurasian Lystrosaurus suggest they are a taxonomically distinct grouping of Lystrosaurus species, which presumably includes L. georgi.
~ By Scott Reid
Sources under the cut
Benoit, J., Angielczyk, K.D., Miyamae, J.A., Manger, P., Fernandez, V. and Rubidge, B., 2018. Evolution of facial innervation in anomodont therapsids (Synapsida): Insights from X‐ray computerized microtomography. Journal of morphology, 279(5), pp.673-701
Botha-Brink, J. (2017). Burrowing in Lystrosaurus: Preadaptation to a postextinction environment? Journal of Vertebrate Paleontology, 37(5), e1365080
Botha-Brink, J., Codron, D., Huttenlocker, A.K., Angielczyk, K.D. and Ruta, M., 2016. Breeding young as a survival strategy during Earth’s greatest mass extinction. Scientific Reports, 6, p.24053
Botha, J. & Smith, R.M.H. (2005). "Lystrosaurus species composition across the Permo–Triassic boundary in the Karoo Basin of South Africa". Lethaia. 40 (2): 125–137
Brink, A. S. (1951). On the genus Lystrosaurus Cope. Transactions of the Royal Society of South Africa, 33(1), 107–120
Broom, R. (1903b). On the structure of the shoulder girdle in Lystrosaurus. Annals of the South African Museum, 4, 139–141
Camp, J. A. 2010. Morphological variation and disparity in Lystrosaurus (Therapsida: Dicynodontia). M.S. thesis, University of Iowa, Iowa City, Iowa, 141 pp
Cluver, M. A. (1971). The cranial morphology of the dicynodont genus Lystrosaurus. Annals of the South African Museum, 56, 155–274
Colbert, E. H. 1974. Lystrosaurus from Antarctica. American Museum Novitates 2535:1-4
Cosgriff, J.W., Hammer, W.R. and Ryan, W.J., 1982. The Pangaean reptile, Lystrosaurus maccaigi, in the Lower Triassic of Antarctica. Journal of Paleontology, pp.371-385
Das, D.P. and Gupta, A., 2012. New cynodont record from the lower Triassic Panchet Formation, Damodar valley. Journal of the Geological Society of India, 79(2), pp.175-180
Grine, F. E., Forster, C. A., Cluver, M. A., & Georgi, J. A. (2006). Cranial variability, ontogeny, and taxonomy of Lystrosaurus from the Karoo Basin of South Africa. In M. T. Carrano, T. J. Gaudin, R. W. Blob & J. R. Wible (Eds.), Amniote paleobiology: Perspectives on the evolution of mammals, birds, and reptiles. (pp. 432–503). Chicago: University of Chicago Press
Groenewald, G. H. (1991). Burrow casts from the Lystrosaurus-Procolophon Assemblage Zone, Karoo Sequence, South Africa. Koedoe, 34(1), 13–22
Gow, C.E. 1999. The Triassic reptile Palacrodon browni Broom, synonymy and a new specimen. Palaeontologia Africana 35: 21–23.
Gubin, Y.M. and Sinitza, S.M., 1993. Triassic terrestrial tetrapods of Mongolia and the geological structure of the Sain-Sar-Bulak locality. The Nonmarine Triassic, 3, pp.169-170
Jasinoski, S.C., Rayfield, E.J. and Chinsamy, A., 2009. Comparative feeding biomechanics of Lystrosaurus and the generalized dicynodont Oudenodon. The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology: Advances in Integrative Anatomy and Evolutionary Biology, 292(6), pp.862-874
Jun, L.I.U., JinLing, L.I. and CHENG, Z., 2002. THE LYSTROSAURUS FOSSILS FROM XINJIANG AND THEIR BEARING ON THE TERRESTRIAL PERMIAN TRIASSIC BOUNDARY. Vertebrata Pal Asiatica, 40(4), pp.267-275
King, G. M., & Cluver, M. A. (1991). The aquatic Lystrosaurus: An alternative lifestyle. Historical Biology, 4, 323–341
Li, J. (1988). Lystrosaurs of Xinjiang, China. Vertebrata PalAsiatica, 26 (4): 241–249
Modesto, S. P., & Botha-Brink, J. (2010). A burrow cast with Lystrosaurus skeletal remains from the Lower Triassic of South Africa. Journal of Vertebrate Paleontology, 25, 274–281
Ray, S. 2005. Lystrosaurus (Therapsida, Dicynodontia) from India: taxonomy, relative growth and cranial dimorphism. Journal of Systematic Palaeontology 3:203–221
Ray, S., Chinsamy, A., & Bandyopadhyay, S. (2005). Lystrosaurus murrayi (Therapsida, Dicynodontia): Bone histology, growth, and lifestyle adaptations. Palaeontology, 48, 1169–1185
Retallack, G.J., 1996. Early Triassic therapsid footprints from the Sydney basin, Australia. Alcheringa, 20(4), pp.301-314
Rozefelds, A.C., Warren, A., Whitfield, A. and Bull, S., 2011. New evidence of large Permo-Triassic dicynodonts (Synapsida) from Australia. Journal of Vertebrate Paleontology, 31(5), pp.1158-1162
Smith RMH, Rubidge BS, van der Walt M. 2011. Therapsid biodiversity patterns and palaeoenvironments of the Karoo Basin, South Africa. In Forerunners of mammals (ed. A ChinsamyTuran), pp. 31 – 64. Bloomington, Indiana: Indiana University Press
Surkov, M.V., Kalandadze, N.N., and Benton, M.J. (June 2005). "Lystrosaurus georgi, a dicynodont from the Lower Triassic of Russia". Journal of Vertebrate Paleontology. 25 (2): 402–413
Taylor, E.L.; Taylor, T.N. (1993). "Fossil tree rings and paleoclimate from the Triassic of Antarctica". In Lucas, S.G. and Morales, M. (eds.) (eds.). The Nonmarine Triassic. Albuquerque: The New Mexico Museum of Natural History and Science Bulletin. pp. 453–455
Viglietti, P.A., Smith, R.M. and Compton, J.S., 2013. Origin and palaeoenvironmental significance of Lystrosaurus bonebeds in the earliest Triassic Karoo Basin, South Africa. Palaeogeography, Palaeoclimatology, Palaeoecology, 392, pp.9-21
Viglietti, P.A.; Smith, R.M.H.; Rubidge, B.S. (2018). "Changing palaeoenvironments and tetrapod populations in the Daptocephalus Assemblage Zone (Karoo Basin, South Africa) indicate early onset of the Permo-Triassic mass extinction". Journal of African Earth Sciences. 138: 102–111
Watson, D. M. S. (1912). The skeleton of Lystrosaurus. Records of the Albany Museum, 2, 287–299
Watson, D. M. S. (1913). The limbs of Lystrosaurus. Geological Magazine, 10(06), 256–258
#lystrosaurus#dicynodont#synapsid#triassic#triassic madness#triassic march madness#prehistoric life#paleontology
188 notes
·
View notes
Text
Lisowicia bojani

By Scott Reid
Etymology: From Lisowice
First Described By: Sulej & Niedźwiedzki, 2019
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Synapsida, Eupelycosauria, Sphenacodontia, Sphenacodontoidea, Therapsida, Eutherapsida, Neotherapsida, Anomodontia, Chainosauria, Dicynodontia, Therochelonia, Bidentalia, Dicynodontoidea, Kannemeyeriiformes, Stahleckeriidae, Placeriinae
Status: Extinct
Time and Place: Approximately between 208 to 215 million years ago, in the late Norian to possibly the earliest Rhaetian of the Late Triassic
Lisowicia is known from the Lipie Śląskie clay−pit near Lisowice, Poland, as well as possibly other coeval sites in Poland
Physical Description: Lisowicia was a very large dicynodont. In fact, with an estimated length of more than 4 metres, a hip height over 2 metres, and an average body weight of 5.88 tons (and possibly up to 7 tons), it was the largest non-mammalian synapsid period, and would be the largest synapsids would ever grow to until the Eocene after the non-avian dinosaurs died out. By and large (ha!), Lisowicia resembles other closely related Triassic dicynodonts like Placerias; it was heavily built with a large barrel-shaped torso and short, stocky legs, a short stubby tail and a large head with a relatively long snout and a tortoise-like beak. Lisowicia was apparently tuskless, like some other Triassic dicynodonts, meaning that it was entirely toothless.
Lisowicia is not only distinctive for its massive size, but also for the design of its limbs. Most dicynodonts had either fully sprawled limbs, or upright hindlimbs and sprawling forelimbs as is the case for larger dicynodonts. Uniquely for dicynodonts, and for any stem-mammal for that matter, Lisowicia had an entirely upright stance with all four limbs being fully erect, holding them directly under its body and swinging them forwards and backwards when moving, like the legs of modern large mammals and dinosaurs. These column-like limbs are no doubt an adaptation for growing to such a massive size, as such limbs are necessary to support large body sizes in land animals, and in the Late Triassic only some sauropodomorph dinosaurs rivalled it in size. This involved not only changes to the orientation of the forelimb, but also changes to the shoulders, shortening of the forearm, and completely rearranging the musculature of the forelimbs in a way no other dicynodont is known to have done. Based on the footprints of other related dicynodonts, the feet of Lisowicia would have been very elephantine in appearance. Like elephants, they would have walked on the tips of the toes with a large fleshy pad underneath the heel to cushion the feet and support its weight. Basically, almost everything about Lisowicia was built to support a massive body size.
The outside appearance of Lisowicia is unknown, and the appearance of most stem-mammals in general is poorly understood. It’s possible it was covered in scales, had naked skin, or even hair, or maybe it had some other unknown structures. If it was anything like large herbivorous dinosaurs and mammals though, it may have been mostly naked anyways.
Diet: Lisowicia was a herbivore, browsing on low and mid-level vegetation using its long snout and toothless beak to crop and chew plants. Remarkably, coprolites attributed to Lisowicia can tell us what they were eating! Lisowicia appears to have mostly preferred eating softer vegetation, along with conifers, and in some cases they even supplemented it with pieces of wood! It’s possible this was a seasonal thing, feeding on tougher, fibrous vegetation during harder times.
Behavior: Like other large herbivores, Lisowicia may have been a herd animal based on fossil sites with multiple individuals preserved together, including adults and juveniles mixed together. Whether this means Lisowicia practised parental care is uncertain though, as it is with most other stem-mammals. Stem-mammal reproduction is poorly understood, but it’s at least probable that they laid eggs, and that includes Lisowicia. Other possible line of evidence for Lisowicia being social could be that they congregated in a specific place to defecate, called communal latrines. Hundreds of coprolites are known from where Lisowicia was first found, and other dicynodonts like Dinodontosaurus are known to have performed similar behaviour, so it’s possible Lisowicia did this too.
Ecosystem: The ecosystem in Late Triassic Poland is rather unusual, as it includes a mix of plants and animals that make it difficult to pin down just how old it really was. Some suggest an older Norian age, others seem more like they’re from younger Rhaetian ecosystems, and it’s still a bit of a mystery to what was going on in this environment compared to the rest of Europe at the time. In any case, the environment where Lisowicia was found was fairly wet and lush, an “everglades-like” swamp dominated by several types of conifers, alongside gingkos, cycads, seed ferns and liverworts. Lisowicia was one of the most common animals in this environment, but it coexisted with a wide variety of other animals. One of the most notable is the predatory archosaur Smok. Smok was the arch predator in this ecosystem, and is known to have directly fed upon Lisowicia, although whether they were actively hunted by it or just scavenged upon is unknown. Smok was certainly capable of crushing even the heavyset bones of Lisowicia, with not only bite marks on its bones attributed to Smok but even broken shards of its bones identified in Smok coprolites! Other reptiles that coexisted with Lisowicia included pterosaurs, dinosauromorphs and predatory theropods similar to Coelophysis or Liliensternus, a small predatory crocodylomorph, and small diapsids, including a sphenodont (a tuatara relative), a possible archosauromorph, and even a possible choristodere. The only other synapsid around was the little mammaliaform cynodont Hallautherium, which was many times smaller than the gigantic Lisowicia. There were also temnospondyl amphibians, including a large predatory capitosaur and a small plagiosaur (one of those very flat temnospondyls with the wide heads), as well as lungfish, coelacanths and even a hybodont shark. Notably, there are no sauropodomorphs known from this ecosystem, even though they were clearly already present in Europe at this time (e.g. Plateosaurus). It’s possible that Lisowicia occupied their role as a giant herbivore in their absence, although whether it had outcompeted them or just happened to live in a habitat without sauropodomorphs and got big is yet another mystery.
Other: In addition to achieving a size and stature amongst synapsids that wouldn’t be seen again until the mammals radiated during the Cenozoic, Lisowicia was also unique for how it grew. While some stem-mammals are known to have grown rapidly as young juveniles, Lisowicia is the only one known so far to never slow down its growth rate at all until adulthood. This type of continuous rapid growth is well known in dinosaurs and mammals, but is unique to Lisowicia amongst dicynodonts. Lisowicia is one of the most unique dicynodonts ever discovered, both as the largest and one of the most derived. It was also sadly one of the last. The cause of its—and all the other Late Triassic dicynodonts’—extinction is unknown, although it was probably a victim of the end-Triassic extinction event like so many other prominent Triassic animals. Late Triassic dicynodonts were once thought to be geographically restricted, left-over relicts from the past that eventually just fizzled away as “more advanced” animals pushed them out. But now we know that they were still globally widespread even in the latest Triassic, including in Europe with advanced forms like Lisowicia. Competition with sauropodomorphs was suspected, but the discovery of the related dicynodont Pentasaurus coexisting with sauropodomorphs in South Africa suggests they were capable of at least living together, and Lisowicia is proof dicynodonts could achieve similarly gigantic proportions. Ultimately, the extinction of the dicynodonts is a mystery, but Lisowicia shows that they were still getting bigger and better than ever, even at the end of their dynasty.
~ By Scott Reid
Sources under the Cut
Bajdek, P., Owocki, K. and Niedźwiedzki, G., 2014. Putative dicynodont coprolites from the Upper Triassic of Poland. Palaeogeography, Palaeoclimatology, Palaeoecology, 411, pp.1-17
Dzik, Jerzy; Sulej, Tomasz; Niedźwiedzki, Grzegorz (2008). "A Dicynodont-Theropod Association in the Latest Triassic of Poland". Acta Palaeontologica Polonica. 53 (4): 733–738
Fiorelli, L.E., Ezcurra, M.D., Hechenleitner, E.M., Argañaraz, E., Taborda, J.R., Trotteyn, M.J., Von Baczko, M.B. and Desojo, J.B., 2013. The oldest known communal latrines provide evidence of gregarism in Triassic megaherbivores. Scientific reports, 3, p.3348
Kowal-Linka, M., Krzemińska, E. and Czupyt, Z., 2019. The youngest detrital zircons from the Upper Triassic Lipie Śląskie (Lisowice) continental deposits (Poland): Implications for the maximum depositional age of the Lisowice bone-bearing horizon. Palaeogeography, palaeoclimatology, palaeoecology, 514, pp.487-501
NIEDŹWIEDZKI, G., GORZELAK, P. and Sulej, T., 2011. Bite traces on dicynodont bones and the early evolution of large terrestrial predators. Lethaia, 44(1), pp.87-92
Romano, Marco; Manucci, Fabio (14 June 2019). "Resizing Lisowicia bojani: volumetric body mass estimate and 3D reconstruction of the giant Late Triassic dicynodont". Historical Biology. 0: 1–6
Qvarnström, Martin; Ahlberg, Per E.; Niedźwiedzki, Grzegorz (2019). "Tyrannosaurid-like osteophagy by a Triassic archosaur". Scientific Reports. 9
Sulej, T., Bronowicz, R., Tałanda, M. and Niedźwiedzki, G., 2010. A new dicynodont–archosaur assemblage from the Late Triassic (Carnian) of Poland. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 101(3-4), pp.261-269
Sulej, T. and Niedźwiedzki, G., 2019. An elephant-sized Late Triassic synapsid with erect limbs. Science, 363(6422), pp.78-80
Świło, M., Niedźwiedzki, G. and Sulej, T., 2013. Mammal-like tooth from the Upper Triassic of Poland. Acta Palaeontologica Polonica, 59(4), pp.815-821
#Lisowicia#Dicynodont#Triassic March Madness#Palaeoblr#Synapsid#Lisowicia bojani#Triassic Madness#Prehistory#Paleontology#Prehistoric life#Mammal-like reptile#Stem mammal
218 notes
·
View notes
Text
Brasilodon

By Ripley Cook
Etymology: Brazilian tooth
First Described By: Bonaparte et al., 2003
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota, Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Synapsida, Eupelycosauria, Metopophora, Haptodontiformes, Sphenacomorpha, Sphenacodontia, Pantherapsida, Sphenacodontoidea, Therapsida, Eutherapsida, Neotherapsida, Cynodontia, Epicynodontia, Eucynodontia, Probainognathia, Chiniquodontoidea, Prozostrodontia, Brasilodontidae
Referred Species: B. tetragonus, B. quadrangularis
Status: Extinct
Time and Place: 227 to 208 million years ago, in the Norian of the Late Triassic.

Take a guess where a synapsid whose name is “Brazilian tooth” was found.
Physical Description: Brasilodon would have looked like a pretty average cynodont in life, because it was. Brasilodon’s skull was wider than deep, as was the case with many cynodonts. Its teeth were distinguished into incisors, canines, and molars. The teeth were small, and the incisors were pointy. The molars were taking on the multicusped shape that modern mammals have, unlike more basal synapsids (which had simple pointy cheek teeth).The teeth were replaced in a polyphyodont manner, where the teeth can theoretically be replaced throughout the animal’s life (unlike most mammals, which get one tooth replacement, and that’s it). Brasilodon’s inner ears indicate a degree of hearing ability close to those of modern mammals, but it lacked the tympanic bone (one of the mammalian ear bones). The rest of the body would have looked rather badgery, with semi-sprawling limbs. Many anatomical details of the postcrania are more similar to those of mammals than other cynodonts. Brasilodon was small, at around 12 cm long, and would have probably been very cute.
Diet: Brasilodon was probably a carnivore. Due to its small size and small, pointy teeth, it may have eaten insects, but its jaw was more resistant to stress than other probainognathians, hinting it may have also taken larger prey.
Behavior: Brasilodon probably had a generalized, adaptable lifestyle. Its forelimbs indicate powerful retractor muscles, but don’t have the same level of robusticity as many burrowing mammals. There are also features of the humerus in common with opossums and primates, and the limbs show a greater degree of flexibility than other probainognathians, which could hint that Brasilodon may have been able to climb. Or… they could just mean nothing; at this size, these adaptations may have been simply to get over irregular, rocky terrain. Nevertheless, burrowing and climbing are both ways to escape potential predators, and climbing could have put Brasilodon in closer proximity to tasty insects.
Ecosystem: Brasilodon was found in the Caturrita Formation of Brazil. In the Triassic, the region was probably fairly arid, with lazy rivers flowing through. There was enough water to support the growth of conifers, at least. This environment was also the home of other cynodonts, such as Brasilitherium, Irajatherium, Minicynodon, and Riograndia, the latter of which was more adapted for burrowing than Brasilodon. Also in this environment lived the sauropodomorph Guaibasaurus, the enigmatic lepidosaur Cargninia, the rhynchocephalian Clevosaurus, and beetles. There was also Faxinalipterus, which was originally described as an early pterosaur, but is probably just some random small reptile.
Other: Brazilodon is one of the very closest relatives of Mammaliaformes (the clade that includes mammals and the very mammaliest synapsids, like Adelobasileus, Morganucodon, and docodonts). Although it has many mammal-like traits, it just wasn’t quite there yet.
~ By Henry Thomas
Sources under the cut
Bonaparte, J.F., Martinelli, A.G., Schultz, C.L. 2005. On the sister-group of mammals: Brasilodon and Brasilitherium (Cynodontia, Probainognathia) from the late Triassic of southern Brazil. Revista Brasileira de Paleontologia 8(1): 25-46.
Guignard, M.L., Martinelli, A.G., Soares, M.B. 2019. The postcranial anatomy of Brasilodon quadrangularis and the acquisition of mammaliaform traits among non-mammaliaform cynodonts. PLoS ONE 14(5): e0216672.
Martinelli, A.G., Bonaparte, J.F. 2010. Postcanine replacement in Brasilodon and Brasilitherium (Cynodontia, Probainognathia) and its bearing in cynodont evolution. In: Calvo, J., Poriri, J., Gonzalez Riga, B., Dos Santos, D. eds. Paleontologia y dinsoaurios desde America Latina. Pp. 179-186.
Salcido, C.J., Rayfield, E.J., Gill, P., Soares, M.B., Martinelli, A.G. 2018. Skull mechanics and functional morpholoy of Brasilodontidae, the sister clade to mammals. Society of Vertebrate Paleontology Annual Meeting, Albuquerque.
Tatarinov, L.P. 2010. On the Origin of the Tympanic Membrane: the Middle Ear of Mammals. Paleontological Journal 44(1): 92-94.
#brasilodon#brasilodontid#probainognathian#cynodont#synapsid#triassic#triassic madness#triassic march madness#prehistoric life#paleontology
152 notes
·
View notes
Text
Morganucodon

By Scott Reid
Etymology: Tooth from Glamorgan (a county in Wales)
First Described By: Kühne, 1949
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Synapsida, Eupelycosauria, Sphenacodontia, Sphenacodontoidea, Therapsida, Eutherapsida, Neotherapsida, Theriodontia, Eutheriodontia, Cynodontia, Probainognathia, Chiniquodontoidea, Prozostrodontia, Mammaliaformes, Morganucodonta, Morganucodontidae
Referred Species: M. watsoni, M. oehleri, M. tardus, M. peyeri, M. heikuopengensis
Status: Extinct
Time and Place: Morganucodon is known from the Kayenta Formation of Arizona, the Forest Marble Formation of England, the Varangeville Conglomerate of France, the Gruhalde Member of the Klettgau Formation of Austria, and the Lufeng Formation of Yunnan, China. Please note that only the Chinese, Austrian, and French locations are of Triassic origin.

Morganucodon lived from the Rhaetian of the Late Triassic to the Bathonian Middle Jurassic of Eurasia, from around 205 million years ago until 167 million years ago.

Physical Description: While Morganucodon itself is not known from many body fossils, its appearance can be inferred from its relatives, such as Megazostrodon, that have been better preserved. Its body would have been elongated and low to the ground, somewhat resembling a shrew. It may have sprawled somewhat with its forelimbs, but its hind legs and hips were much closer to those of mammals. Morganucodon’s skull was long and flat on top. It had two types of teeth, premolars and molars, similar to later mammals. The mammaliaform also replaced its teeth once in its lifetime: it had “baby” (or “milk”) teeth and “adult” teeth, like its modern relatives.
Diet: Morganucodon was almost certainly insectivorous. Study of its teeth showed that its jaws could stand up to more stress and that its bite was relatively forceful, allowing members of the genus to hunt “tougher” prey items than its contemporaries. It may have eaten beetles, which are known from its environment.
Behavior: These animals were probably nocturnal, as evidenced by heightened senses of hearing and smell, as well as comparisons with similar modern animals.
Ecosystem: Due to its wide distribution, Morganucodon would likely have inhabited a range of environments. The first fossils were discovered in what were once fissures on an early Jurassic or late Triassic limestone island, St. Bride’s Island. It was a small landmass of about 20 square km (7.7 square mi) with a karst environment of sinkholes and caves. It would also have been warm, supporting plants such as clubmosses, bennetites, ferns, and cycads. Morganucodon would have shared its environment with other mammaliaforms, such as Kuehneotherium, and as such likely experienced niche partitioning (different species occupying different niches in ecosystems, which reduces competition). Other animal species known from the same areas include ancient reticulated beetles, the sphenodontian Gephyrosaurus, and another mammal relative, Oligokyphus.
The Swiss fossils were unearthed in marine deposits, though it is likely that it was washed out to sea. Other terrestrial animals known from these beds include the pterosaur Peteinosaurus, the trilophosaur Variodens, a few rhynchocephalians, and several species of cynodonts. The environment of the French specimens is less well-documented, but likely similar, with therapsids, temnospondyl amphibians, pterosaurs, and phytosaurs all being documented.
Other: This small mammaliaform has helped to illuminate the evolution of mammalian ears. Our lower jaws, and those of our modern mammalian cousins, are unique in that they are made up of only one bone: the mandible. In other animals, this structure is made up of multiple different bones, which are used to help with chewing. Somewhere along the line in mammal evolution, most of our lower jaw bones shrunk in size and moved to our ears, enabling us to have a complex system of hearing. Morganucodon represents an intermediate step in this process: it still had many of the jaw bones of more basal amniotes, albeit much more reduced.
It would also likely have been warm-blooded, as evidenced by its secondary palate. This feature of the skeleton separates the nasal cavity from the mouth, allowing animals with it to quickly breathe while using their mouths for feeding. Morganucodon also had a bone structure that suggested rapid growth. Other shared traits with modern mammals include the likely presence of mammary glands and a relatively large brain.
~ By Jasper Barnes
Sources under the Cut
Benton, Michael J. (2015). Vertebrate Palaeontology (fourth edition). Wiley Blackwell.
Butler, Percy M., Sigogneau-Russell, Denise (2016). Diversity of triconodonts in the Middle Jurassic of Great Britain. Palaeontologia Polonica 67, 35–65.
Evans, S. E. (1992). Small reptiles and amphibians from the Forest Marble (Middle Jurassic) of Dorset. Proceedings of the Dorset Natural History and Archaeological Society 113:201-202
Fraser, Nicholas (2006). Dawn of the dinosaurs: life in the Triassic. Indiana University Press.
Gill, Pamela G., Purnell, Mark A., Crumpton, Nick., Robson Brown, Kate, Gostling, Neil J., Stampanoni, M., and Rayfield, Emily J. (2014). Dietary specializations and diversity in feeding ecology of the earliest stem mammals. Nature 512, 303–305
Godefroit, P. (1997). Reptilian, therapsid and mammalian teeth from the Upper Triassic of Varangéville (northeastern France). Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre 67:83-102
Peyer, B. (1956). Über Zähne von Haramiyiden, von Triconodonten und von wahrscheinlich synapsiden Reptilien aus dem Rhät von Hallau, Kt. Schaffhausen, Schweiz [On the teeth of haramiyids, triconodonts, and probable synapsid reptiles from the Rhaetic of Hallau, Kanton Schaffhausen, Switzerland]. Schweizerische Paleontologische Abhandlungen 72:1-72
Walter G. Kühne, "On a Triconodont tooth of a new pattern from a Fissure-filling in South Glamorgan", Proceedings of the Zoological Society of London, volume 119 (1949–1950) pages 345–350
#Morganucodon#Mammaliaform#Mammal#Triassic#Palaeoblr#Triassic Madness#Triassic March Madness#Morganucodon watsoni#Morganucodon oehleri#Morganucodon tardus#Morganucodon peyeri#Morganucodon heikuopengensis#Prehistoric life#Prehistory#Paleontology
178 notes
·
View notes
Last Seen Blogs




