#* plumule / study.
Note
hey what are your pronouns
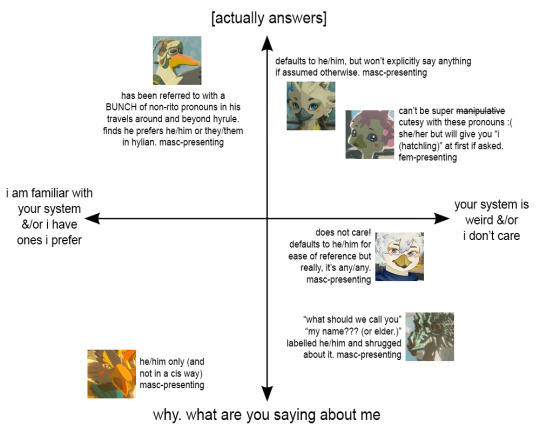
funny you should ask!
transcripts, from top ("[actually answers]") to bottom ("why. what are you saying about me"):
penn: has been referred to with a BUNCH of non-rito pronouns in his travels around and beyond hyrule. finds he prefers he/him or they/them in hylian. masc-presenting
tulin: defaults to he/him, but won't explicitly say anything if assumed otherwise. masc-presenting
molli: can't be super manipulative cutesy with these pronouns :( she/her but will give you "i (hatchling)" at first if asked. fem-presenting
kido: does not care! defaults to he/him for ease of reference but really, it's any/any. masc-presenting
dineli: "what should we call you" -> "my name??? (or elder.)" labelled he/him and shrugged about it. masc-presenting
revali: he/him only (and not in a cis way). masc-presenting
#quillheel#* roosting / ooc.#wow thank you for this question i definitely did not ask to have sent in#asfjdlksg no but really thank you captain!! <3#i have yet to figure out a way to satisfactorily describe the ri.tos' view on gender (as interpreted by me)#because what they have ingame is like uhhhh i'm Not Here For Those Implications. Sorry!#BUT i need y'all to know i only use the pronouns i use for my birdies when writing because it is the simplest and easiest#interpretation of what they'd actually use. and also the closest to what they'd most (< for lack of better words) agree with#even rev.ali who i use 'not [...] cis' with is like....that's not the term they'd use but it's about the same concept slkfjld#the kinship terms system is my best worded post on the topic so you'll have to excuse me for plugging it again!#* arrowhead / study.#* plumule / study.#* powder down / study.#* field notes / study.#* wisps / study.#* vanes / study.
11 notes
·
View notes
Text
Lupine Publishers|Effect of Interaction Between Ag Nanoparticles and Salinity on Germination Stages of Lathyrus Sativus L.
Abstract
The aim of the study was to effect of interaction between Ag nanoparticles and salinity on Germination Stages of Lathyrus Sativus L. Treatments included in the study were viz. To 3 levels of salinity (0 as control, 8 and 16 dS/m NaCl), 8 and 16 dS/m and four levels of silver nanoparticles (0, 5, 10 and 15 ppm) on grass pea seed were tested. An experiment was conducted to evaluate the effects of silver nanoparticles (AgNPs), on the seed germination factors, root and shoot length (RL and SL) and proline content of grass pea Survival under Salinity Levels. Results showed a significant reduction in growth and development indices due to the salinity stress. The salt stress impaired the germination factors of grass pea seedlings. The application of Ag in combination improved the germination percentage, shoot and root length, seedling fresh weight and seedling dry weight and seedling dry contents of grass pea seedlings under stressed conditions. The results suggest that Ag nanoparticles enhancement may be important for osmotic adjustment in grass pea under salinity stress and application of Ag mitigated the adverse effect of salinity and toxic effects of salinity stress on grass pea seedlings.
Keywords: Ag nanoparticles; Salinity; Germination Stages; Grass Pea; Lathyrus Sativus L.
Introduction
High salinity is a common abiotic stress factor that causes a significant reduction in growth. Germination and seedling growth are reduced in saline soils with varying responses for species and cultivars [1]. Soil saltiness may impact the germination of seeds either by causing an osmotic potential outside to the seed averting water uptake, or the poisonous effects of Na+ and Cl− ions on germinating seed [2]. Salt and osmotic stresses are responsible for both inhibition or delayed seed germination and seedling establishment [3]. The majority of our present-day crops are adversely affected by salinity stress [4]. NaCl causes extensive oxidative damage in different legumes, resulting in significant reduction of different growth parameters, seed nutritional quality, and nodulation [5,6]. To mitigate and repair damages triggered by oxidative stress, plants evolved a series of both enzymatic as well as a non-enzymatic antioxidant defense mechanism. Ascorbate and carotenoids are two important non-enzymatic defenses against salinity, whereas proline is the most debated osmoregulatory substances under stress [7].
Lathyrus Sativus L. (Grass pea) is an annual pulse crop belonging to the Fabaceae family and Vicieae tribe [8]. Grass pea has a long history in agriculture. The crop is an excellent fodder with its reliable yield and high protein content. This plant is also commonly grown for animal feed and as forage. The grass pea is endowed with many properties that combine to make it an attractive food crop in drought-stricken, rain-fed areas where soil quality is poor and extreme environmental conditions prevail [9]. Despite its tolerance to drought it is not affected by excessive rainfall and can be grown on land subject to flooding [10,11]. Compared to other legumes, it is also resistant to many insect pests [12-15]. Nanoparticles (NPs) are wide class of materials that include particulate substances, which have one dimension less than 100 nm at least [16]. The importance of these materials realized when researchers found that size can influence the physiochemical properties of a substance e.g. the optical properties [17]. NPs with different composition, size, and concentration, physical/ chemical properties have been reported to influence growth and development of various plant species with both positive and negative effects [18]. Silver nanoparticles have been implicated in agriculture for improving crops. There are many reports indicating that appropriate concentrations of AgNPs play an important role in plant growth [19,20]. The application of Nano silver during germination process may enhance germination traits, plant growth and resistance to salinity conditions in basil seedlings [21]. The use of Silver Nanoparticle on Fenugreek Seed Germination under Salinity Levels is a recent practice studied [22]. Nanomaterials have also been used for various fundamental and practical applications [23]. Although the potential of AgNPs in improving salinity resistance has been reported in several plant species [24,25], its role in the alleviation of salinity effect and related mechanisms is still unknown. Therefore, the main objective of this work was to study the effect of Silver Nanoparticles on salt tolerance in Lathyrus Sativus L.
Material and Methods
In order to investigate salinity stress on Lathyrus Sativus L. germination indices, an experiment was carried out in Iran from April to Juan 2017 at Ferdowsi University of Mashhad, to creation salinity, sodium chloride at the levels of 8 dS/m, 15 dS/m and 0 (as control), four levels of silver nanoparticles (0, 5, 10 and 15 ppm) on Grass pea were tested. The Ag NPs were obtained from US Research Nanomaterial’s, Inc. Transmission electron microscopy (ТЕМ) images of silver nanoparticles with diameters of 20 nm, shown in Figure 1. Seeds of Lathyrus Sativus L. where from seed bank of Research Center for Plant Sciences, Ferdowsi University of Mashhad. These all were washed with deionized water. Seeds were sterilized in a 5% sodium hypochlorite solution for 10 minutes [26], rinsed through with deionized water several times. Their germination was conducted on water porous paper support in Petri dishes (25 seed per dish) at the controlled temperature of 25 ± 1°C. After labeling the Petri dishes, seed were established between two Whatman No. 2 in Petri dishes. Silver nanoparticles in different concentration silver nanoparticles (0, 5, 10 and 15ppm) were prepared directly in deionized water and dispersed by ultrasonic vibration for one hour. Each concentration was prepared in three replicates. Every other day supply with 0.5 ml silver nanoparticles per every test plantlet was carried out for 21 days along with control. Germination counts were recorded at 2 days’ intervals for 21 days after sowing and the seedlings were allowed to grow. The germination percentages of the seeds were finally determined for each of the treatments. After 21 days of growth, the shoot and root lengths were long enough to measure using a ruler. The controlled sets for germinations were also carried out at the same time along with treated seeds (Figure 2).
Figure 1: Silver Nanopowder, Coated with ~0.2wt% PVP (Poly Viny Pyrrolidone) surfactant for low oxygen content and easy dispersing. True density: 10.5 g/cm3 Purity: 99.99% APS: 20 nm SSA: ~18-22 m2/g Color: black, Morphology: spherical.
Figure 2: Effect of Ag Nanoparticles on Germination Stages of Lathyrus Sativus L. in Salinity level( 8 dS/m NaCl).
Parameters Measured in this Study were:
A. Germination Stages
Total germination percentage (GT) was calculated as Gt = (n/N ×100), where n = total number of germinated seeds (normal and abnormal) at the end of the experiment and N = total number of seeds used for the germination test.
B. Germination Speed Index (GSI)
Conducted concomitantly with the germination test, with a daily calculation of the number of seeds that presented protrusion of primary root with length ≥2 millimeter, continuously at the same time amid the trial. The germination speed index was calculated by Maguire formula [27]: aguire formula (1962):
Where:
GSI = seedlings’ germination speed index;
G = number of seeds germinated each day;
N = number of days elapsed from the seeding until the last count.
Root and Shoot Length
Root length was taken from the point below the hypocotyls to the end of the tip of the root. Shoot length was measured from the base of the root- hypocotyl transition zone up to the base of the cotyledons. The root and shoot length were measured with the help of a thread and scale.
Seedling Vigour Index
The seedling vigor index was determined by using the formula given by Abdul baki and Anderson [28].
Fresh and Dry mass
The fresh mass was quantified through weighing on precision scale, and the dry mass was determined through weighing on a precision scale after permanence of the material in a kiln with air forced circulation, at a temperature of 70°C, until indelible weight. At the ending of the experiment, At the end of the experiment, radical and plumule length and fresh weight measured. Plants were placed in the oven at 70°C for 48 h and weighted with sensitive scale.
Proline Contents
Proline was determined spectrophotometrically following the ninhydrin method described, using L-proline as a standard [29]. Approximately 300 mg of dry tissue was homogenized in 10ml of 3% (w/v) aqueous sulphosalicylic acid and filtered. To 2ml of the filtrate, 2ml of acid ninhydrin was added, followed by the addition of 2ml of glacial acetic acid and boiling for 60 min. The mixture was extracted with toluene, and the free proline was quantified spectrophotometrically at 520nm from the organic phase using a spectrophotometer. Statistical analysis each treatment was conducted, and the results were presented as mean ± SD (standard deviation). The results were analyzed by one-way ANOVA with used Minitab Version 16.
Results and Discussion
The present study showed clearly that salinity had a negative effect on the yield and its components of grass pea. It is well known that seed germination provides a suitable foundation for plant growth, development, and yield [30]. Increased salt concentration caused a decrease in germination percent (Table 1). Seed germination decreased as the doses increased. The Strong reduction in germination (-47%) was observed mainly at the highest level of salt concentration as compared to control treatment. Delayed germination causes increased irrigation cost and irregular and weak seedling growth in the establishment of legume crops. Relevant results were reported by Gunjaca and Sarcevic [31] and Almansouri et al. [32]. They reported that increasing osmotic potential decreased water uptake and slow down germination time. The average time of germination increases with increasing levels of salinity. In view of mean germination time, there was a considerable increase in this character at 0 (as control), 8 and 16 DS/m salinity levels as compared to the others. Emergence was significantly affected by salinity levels. Moreover, many researchers have reported developmental delay of seed germination at high salinity [33]. The germination rate decreased as salt concentration increased to a 16 dS/m and delayed for the high salt dosage (Table 1). Since higher salinity limited water absorption, it has prevented nutrient assimilation, as a result, germination rate declined with increasing salinity. The findings from this study were like to the findings of Kaydan and Yagmur [34] and Akhtar and Hussain [35].
Table 1: The interaction effect of NaCl and AgNPs on Germination Speed Index.
Table 2: Analysis of variance of the measured traits.
Shoot fresh weight was significantly influenced (P<0.05) by salinity levels. The highest shoot fresh weight was obtained from 0dSm salinity level while the lowest weight was at 16dSm. Shoot fresh weight significantly decreased as salinity level increased above 8dSm (Table 2). Salinity stress significantly (P<0.05) affected shoot dry weight as the salt concentration dosage increased. Shoot dry weight significantly decreased in salt levels over 8dSm. When the salinity level was raised above, the proline content increased in grass pea. Culturing excised roots has demonstrated to be a really great test show for the early detection of tolerance to abiotic stresses such as saltiness [36-38].
Proline was studied in numerous works dealing with plant selection against abiotic stresses such as dry and salinity [39,40], and it may play a defensive part against the osmotic potential produced by salt [41,42]. The proline substance of the expanded with the NaCl concentration of the culture medium. At 16 dS/m NaCl, the proline concentration appeared a huge increment in reaction to salt stress, although the activity of the roots at this concentration was negligible, with no grateful longitudinal development. Proline, which happens broadly in higher plants and collects in bigger sums than other amino acids [43], regulates the aggregation of useable N. Proline collection normally occurs within the cytosol where it contributes significantly to the cytoplasmic osmotic alteration [44]. It is osmotically very active and contributes to membrane stability and mitigates the impact of on NaCl cell membrane disturbance [45]. In the present experiment application of Ag NPs enhanced seed potential by increasing the characteristics of seed germination (Tables 1 & 2). The results showed that the impact of Ag NPs was significant on germination percentage in P≤0. 05. The results about of this test appeared that utilization of Ag NPs nanoparticles can increment the germination in grass pea. Seed germination results indicate that Ag Nanoparticles at their lower concentrations advanced seed germination and early seedling growth in grass pea, anyway at higher concentration showed slight antagonistic impacts. Parameters of seed germination were expanded with increasing levels of Ag NPs up to 10 ppm. Among the treatments, application of 10 ppm of Ag NPs proved best by giving the highest values for percent seed germination, germination rate and germination mean time. It is well watched that the exogenous application of Ag NPs decreased the reduction of germination resulted from salt treatments. In the interim, the control treatments of salt and Ag nanoparticles gave the tallest plants contrasted with the other studied treatments. Darvishzadeh et al. [21] found that the utilization of Ag Nano particles at the concentration of 40 mg.kg-1 prompted the increases in germination percentage and improved the resistance to salinity conditions in basil. The proline content increased with increasing severity of salinity stress. Additionally, proline content significantly (P.0.01) increased when silver nanoparticles were applied in connected in serious saline stress in comparison without silver nanoparticles (Figure 3).
Figure 3: Effect of interaction between ag nanoparticles and salinity on germination stages of Lathyrus Sativus L. a - Main effects plot for Root length (mm); b - Main effects plot for dry weight of shoot and root (gr); c - Interaction Plot for fresh weight of shoot and root (gr); d - Interaction Plot for dry weight of shoot and root (gr); e - Interaction Plot for Proline (mg/gr).
Conclusion
Salt stress through enhancement of osmotic pressure leads to the decrease of germination percentage, germination rate, germination index and an increment in mean germination time of Lathyrus sativus seeds. For overcoming the negative impacts of salinity on the plant growth and yield can be to attempt to new strategies. The dry and fresh weight of seedlings diminished as seedling length declined with increasing salinity levels since root number, shoot number, root length and shoot length decreased essentially. Results demonstrate that Ag NPs at lower concentration enhances seed germination, promptness index, and seedling growth. The positive effect of Ag on physiological properties was in conditions that the plant grew under salt stress was more increasingly exceptional in examination with the conditions that plant grown under normal conditions. The results of this study showed that Ag can be involved in the metabolic or physiological activity in higher plants exposed to abiotic stresses.
Acknowledgment
I would like to thank the Research Center for Plant Sciences, and Dr. Jafar Nabati for providing the necessary facilities.

#Lupinepublishers#Lupine Publishers Group#open access journals#Earth and Environment Journals#ecology
2 notes
·
View notes
Text
Reduction in Substrate Moisture Content Reduce Final Yield of Wheat - BJSTR Journal

Reduction in Substrate Moisture Content Reduce Final Yield of Wheat by Syed Arshad Karim and Imran* in Biomedical Journal of Scientific & Technical Research
https://biomedres.us/fulltexts/BJSTR.MS.ID.002656.php
Wheat (Triticum aestivum L.) is an important crop of Pakistan. In Khyber Pakhtunkwa, wheat is grown on almost 67% cropped area. Its yield is subjected to availability of rainfall. Delay in rain caused severe yield reduction by early drought stress. Therefore, this experiment was conducted to compare new wheat varieties in relation to reduction in substrate moisture content to find out drought tolerant variety that could withstand the early drought stress. The experiment was conducted in Completely Randomized Design (CRD) in Agronomy Lab, Department of agronomy, the University of Agriculture Peshawar in controlled environment in 2016. Five different wheat varieties seeds were compared in early growth establishment in three repeats with three different moisture level,
a) Low moisture (6%),
b) Medium moisture (12%),
c) High moisture (18%).
The growth substrate silt was used in specially designed glass boxes box of size (23 x 16 x 4 cm). All the other environmental conditions were kept similar for each variety and its replicates. Data was recoded on different growth parameters such as germination percentage, root number, plumule length, dry weight of shoot and root, and length of seminal root 1, 2, and 3. The study confirmed that almost all varieties can withstand at low moisture but significant reduction in different parameters was observed with reduction in moisture content of growth media that could ultimately affect the yield of wheat.
For more articles on Journals on Medical Drug and Therapeutics please click here
bjstr
Follow on Twitter : https://twitter.com/Biomedres01
Follow on Blogger :https://biomedres01.blogspot.com/
Like Our Pins On : https://www.pinterest.com/biomedres/
#Journals on Biomedical Intervention#Journals on Biomedical Imaging#Journals on Medical Informatics#Journals on Cancer Medicine#Journal on Medical Genetics
0 notes
Text
Dictionary (pt.cclx)
Words taken from This Book Was a Tree by Marcie Chambers Cuff:
Demodex (n.)
a genus of tiny mites that live in or near hair follicles of mammals.
epidemiology (n.)
the study of the incidence and distribution of diseases, and of their control and prevention.
plumule (n.)
the rudimentary shoot or stem of an embryo plant.
pluton (n.)
an intrusive body of plutonic rock, especially a large one.
mascarpone (n.)
a soft mild Italian cream cheese.
radicle (n.)
the part of a plant embryo that develops into the primary root; a rootlet.
alfalfa (n.)
a leguminous plant, Medicago sativa, with clover-like leaves and flowers used for fodder.
camphor (n.)
a white translucent crystalline volatile substance with an aromatic smell and bitter taste, used to make celluloid and in medicine. Chemical formula: C10H16O.
hypocotyl (n.)
the part of the stem of an embryo plant beneath the stalks of the seed leaves or cotyledons and directly above the root.
witch hazel (n.)
any North American shrub of the genus Hamamelis, with bark yielding an astringent lotion.
1 note
·
View note
Text
Defense Mechanisms of Plants to Insect Pests: From Morphological to Biochemical Approach

Abstract
The plants defend themselves against insect herbivores through utilizing the combination of direct defense traits and indirect defense approaches. Direct defense is involved both physical and chemical barriers which synergistically obstruct insect herbivore's growth, development, reproduction, etc. The indirect defense approach has no direct impact on insect herbivores but suppress pests by releasing volatile compounds that attract natural enemies of the herbivores. Plant defense against insect herbivores is just one of multiple layers of interactions. Together with plants, these players are involved in complex interaction networks. To elucidate these fascinating interactions biochemical, ecological as well as molecular studies, and combinations thereof, are required.
Keywords: Insects; Defense mechanisms; Biochemical defense; Secondary metabolites; Induced defense mechanisms
Abbreviations: PIs: Proteinase Inhibitors; BXs: Benzoxazinoids; HIPVs: Herbivore Induced Plant Volatiles; CDPK: Calcium-Dependent Protein Kinases
Introduction
Insects are one of the dominant forms of life in terms of the number of species and of individuals. Plants can have different types of interactions with insects, such as antagonistic interactions with herbivores and mutualistic interactions with carnivorous and pollinating insects. Plants can defend themselves against insects by employing a 'do-it- yourself' strategy and/or by enlisting 'bodyguards' that attack herbivores. These plant strategies can be present constitutively or they can be induced by herbivory. Inducible defenses result in variable plant phenotypes and consequently in variable types of interactions with insects [1].
Plants respond to herbivore through various morphological, biochemical, and molecular mechanisms to counter/offset the effects of herbivore attack. According to Oerke [2], each year there is a huge crop yield loss by different insect pests around the world. Therefore, understanding the defensive systems or mechanisms of plants enables development of resistant crops or pest management systems reducing the need of hazardous pesticides and supporting safer crop production. Another positive effect would be a reduction of the development of pesticide resistant pest strains.
Insect herbivores have traditionally been divided into generalists (polyphagous) that feed on several hosts from different plant families, or specialists (monophagous and oligophagous), which feed on one or a few plant types from the same family. The generalists tolerate a wide array of defenses present in most plants, while they cannot feed on certain plants that have evolved more unique defense mechanisms. Specialists, on the other hand, use a specific range of host plants releasing defense compounds that at the same time may function as feeding stimulants and provide ovipositioning cues [3].
Pegadaraju et al [4] stated that, the defensive mechanisms in plants operate at different levels. They vary from external defenses like thorns to complicated chemical responses leading to poisoning of the attacker. To overcome the insect attack, plants produce specialized morphological structures or secondary metabolites and proteins that have toxic, repellent, and/or anti nutritional effects on the insect pests. In addition, plants also release volatile organic compounds that attract the natural enemies of the herbivores [5].
Resistance factors for direct plant defense against herbivorous insects comprise plant traits that negatively affect insect preference (host plant selection, oviposition, feeding behavior) or performance (growth rate, development, reproductive success) resulting in increased plant fitness in a hostile environment. Such traits include morphological features for physical defense, like thorns, spines, and trichomes, epicuticular wax films and wax crystals, tissue toughness, as well as secretory structures and conduits for latices or resins. They also include compounds for chemical defense, like secondary metabolites, digestibility reducing proteins, and anti nutritive enzymes. All these traits may be expressed constitutively as preformed resistance factors, or they may be inducible and deployed only after attack by insect herbivores. The induction of defensive traits is not restricted to the site of attack but extends to non-infested healthy parts of the plants. The systemic nature of plant responses to herbivore attack necessitates a long-distance signaling system capable of generating, transporting, and interpreting alarm signals produced at the plant-herbivore interface. Much of the research on the signaling events triggered by herbivore has focused on tomato and other solanaceous plants. In this model system, the peptide system in acts at or near the wound site to amplify the production of jasmonic acid. Jasmonic acid or its metabolites serve as phloem-mobile long-distance signals, and induce the expression of defense genes in distal parts of the plant [6].
Host Plant Defenses against Insects
Plants respond to insect attack through an intricate and dynamic defense system that includes structural barriers, toxic chemicals, and attraction of natural enemies of the target pests. Both defense mechanisms (direct and indirect) may be present constitutively or induced after damage by the insects. Induced response in plants is one of the important components of insect pest control in agriculture, and has been exploited for regulation of insect herbivore population.
Induced defenses make the plants phenotypically plastic, and thereby, decrease the chances of the attacking insects to adapt to the induced chemicals. Changes in defensive constituents of a plant on account of insect attack develop unpredictability in the plant environment for insect herbivores, which in turn, affects the fitness and behavior of the insects. If induced response occurs very early, it is of great benefit to the plant, and reduces the subsequent herbivore and pathogen attack, besides improving overall fitness of the plant. Plants with high variability in defensive chemicals exhibit a better defense compared with those with moderate variability [7].
Direct defenses
Plant structural traits such as leaf surface wax, thorns or trichomes, and cell wall thickness/and lignification form the first physical barrier to feeding by the insects, and the secondary metabolites such act as toxins and also affect growth, development, and digestibility reducers form the next barriers that defend the plant from subsequent attack. Moreover, synergistic effect among different defensive components enhances the defensive system of plants against the insects' invaders. For example, in tomato, alkaloids, phenolics, Proteinase Inhibitors (PIs), and the oxidative enzymes when ingested separately result in a reduced affect, but act together in a synergistic manner, affecting the insect during ingestion, digestion and metabolism. In a wild tobacco (Nicotiana attenuate), trypsin proteinase inhibitors and nicotine expression, contributed synergistically to the defensive response against beet armyworm (Spodoptera exigua) [7].
Morphological features for physical defense
Insect herbivores from all feeding guilds must make contact with the plant surface in order to establish themselves on the host plant. It is therefore not surprising that physical and chemical features of the plant surface are important determinants of resistance. All plant parts offer some sort of resistance against herbivory. They range from tissue hardness to highly complex glandular trichomes and spines. Epicuticular wax films and crystals cover the cuticle of most vascular plants. In addition to their important role in desiccation tolerance, they also increase slipperiness, which impedes the ability of many non-specialized insects to populate leaf surfaces. The physical properties of the wax layer as well as its chemical composition are important factors of preformed resistance [6].
Based on different findings, plant structures are the first line of defense against insect pests, and play an important role in host plant resistance to insects. The first line of plant defense against insect pests is the erection of a physical barrier either through the formation of a waxy cuticle, and/or the development of spines, setae, and trichomes. Structural defenses includes morphological and anatomical traits that confer a fitness advantage to the plant by directly deterring the herbivores/ insects from feeding, and range from prominent protrubances on a plant to microscopic changes in cell wall thickness as a result of lignification and suberization. Structural traits such as spines and thorns (spinescence), trichomes (pubescence), toughened or hardened leaves (sclerophylly), incorporation of granular minerals into plant tissues, and divaricated branching (shoots with wiry stems produced at wide axillary angles) play a leading role in plant protection against insect pests. Sclerophylly refers to the hardened leaves, and plays an active role in plant defense against herbivores by reducing the palatability and digestibility of the tissues, thereby, reducing the herbivore damage.
Spinescence includes plant structures such as spines, thorns and prickles. It has been reported to defend the plants against many insects. Pubescence consists of the layer of hairs (trichomes) extending from the epidermis of the above ground plant parts including stem, leaves, and even fruits, and occur in several forms such as straight, spiral, stellate, hooked, and glandular. Chamarthi et al. [8] reported that leaf glossiness, plumule and leaf sheath pigmentation were responsible for shoot fly (Atherigona soccata resistance in sorghum Sorghum bicolor (L. Moench).
Trichomes
The plant epidermis is often covered by outgrowths called trichomes. They are found in all major groups of terrestrial plants. They originate from epidermal tissue and then develop and differentiate to produce hair-like structures [9]. Trichomes play an imperative role in plant defense against many insect pests and involve both toxic and deterrent effects. Trichomes density negatively affects the ovipositional behavior, feeding and larval nutrition of insect pests. In addition, dense trichomes affect the insect mechanically, and interfere with the movement of insects and other arthropods on the plant surface, thereby, reducing their access to leaf epidermis. These can be, straight, spiral, hooked, branched, or un-branched and can be glandular or non-glandular. Glandular trichomes secrete secondary metabolites including flavonoids, terpenoids, and alkaloids that can be poisonous, repellent, or trap insects and other organisms, thus forming a combination of structural and chemical defense.
Induction of trichomes in response to insect damage has been reported in many plants [10]. This increase in trichomes density in response to damage can only be observed in leaves developing during or subsequent to insect attack, since the density of trichomes of existing leaves does not change. A given authors reported that damage by adult leaf beetles (Phratora vulgatissima) in Salix cinerea plant induced higher trichome density in the new leaves developing thereafter. Likewise, increase in trichome density in S. cinera in response to coleopteran damage has also been reported. Increase in trichome density after insect damage has also been reported in Lepidium virginicum L. and Raphanus raphanistrum L. In black mustard, trichomes density and glucosinolate levels were elevated after feeding by small white butterfly (Pieris rapae). Furthermore, change in relative proportion of glandular and non-glandular trichomes is also induced by insect.
Leaf and root toughness and quantity
Leaf toughness interferes with the penetration of plant tissues by mouthparts of piercing-sucking insects and increase mandibular wear in biting-chewing herbivores [11]. The cell walls of leaves are also reinforced during feeding through the use of different macromolecules, such as lignin, cellulose, suberin and callose, together with small organic molecules, such as phenolics, and even inorganic silica particles. Roots eaten by insect herbivores exhibit extensive regrowth, both in density, as seen in Trifolium repens eaten by Sitona lepidus (clover root weevil), and in quantity, as observed in Medicago sativa (alfalfa) attacked by clover weevil (Sitona hispidulus). The former might be caused by additional lignification that could increase the toughness of the roots. In addition, genotypes with long fine roots suffered less from herbivory compared to genotypes with short and thick roots.
Secondary metabolites for chemical defense of plants
Plants produce a large and diverse array of organic compounds that appear to have no direct functions in growth and development i.e. they have no generally recognized roles in the process of photosynthesis, respiration, solute transport, translocation, nutrient assimilation and differentiation. These compounds or chemicals play a significant role in direct defense impair herbivore performance by one of two general mechanisms: these chemicals may reduce the nutritional value of plant food, or they may act as feeding deterrents or toxins. There has been considerable debate as to which of these two strategies is more important for host plant selection and insect resistance. An important part of this debate concerns the extent to which variation in the levels of primary and secondary metabolites has evolved as a plant defense [12]. Plant primary metabolism, which is shared with insects and other living organisms, provides carbohydrates, amino acids, and lipids as essential nutrients for the insect.
Secondary metabolites are the compounds that do not affect the normal growth and development of a plant, but reduce the palatability of the plant tissues in which they are produced. The defensive (secondary) metabolites can be either constitutive stored as inactive forms or induced in response to the insect or microbe attack. The former are known as phytoanticipins and the latter as phytoalexins (antimicrobial compounds synthesized by plants that accumulate rapidly at areas of pathogen infection). The phytoanticipins are mainly activated by p-glucosidase during herbivory, which in turn mediate the release of various biocidal aglycone metabolites. The classic examples of phytoanticipins are glucosinolates that are hydrolyzed by myrosinases (endogenous p-thioglucoside glucohydrolases) during tissue disruption. Other phytoanticipins include Benzoxazinoids (BXs), which are widely distributed among Gramineae. Hydrolyzation of BX-glucosides by plastid-targeted p-glucosidases during tissue damage leads to the production of biocidal aglycone BXs, which play an important role in plant defense against insects. Phytoalexins include isoflavonoids, terpenoids, alkaloids, etc., that influence the performance and survival of the insects. The secondary metabolites not only defend the plants from different stresses, but also increase the fitness of the plants. It has been reported that maize to corn earworm, Helicoverpa zea is mainly due to the presence of the secondary metabolites C-glycosyl flavone maysin and the phenylpropanoid product, chlorogenic acid. Compound, 4, 4- dimethyl cyclooctene has been found to be responsible for shoot fly resistance in sorghum [8].
Study on secondary metabolites could lead to the identification of new signaling molecules involved in plant resistance against insect pests. Ultimately genes and enzymes involved in the biosynthesis of these metabolites could be identified. Some of the secondary metabolites in plant defense will be the following.
Plant phenolic compounds
Among the secondary metabolites, plant phenols constitute one of the most common and widespread group of defensive compounds, which play a major role in host plant resistance against insects. Phenols act as a defensive mechanism not only against insects, but also against microorganisms and competing plants.
Lignin, a phenolic heteropolymer plays a central role in plant defense against insects and pathogens. It limits the entry of pathogens by blocking physically or increasing the leaf toughness that reduces the feeding by insects, and also decreases the nutritional content of the leaf. Lignin synthesis has been found to be induced by insect or pathogen attack and its rapid deposition reduce further growth of the pathogen or insect fecundity.
Plant defensive proteins
Ecologically, in insect-plant interaction, interrelationship between two is important for the survival of the both. Insects always look for a true and healthy host plant that can provide them proper food and could be suitable for mating, oviposition and also provides food for the offspring's. The nutritional requirements of insects are similar to other animals, and any imbalance in digestion and utilization of plant proteins by the insects' results in drastic effects on insect physiology. Alteration of gene expression under stress including insect attack leads to qualitative and quantitative changes in proteins, which in turn play an important role in signal transduction, and oxidative defense. Many plant proteins ingested by insects are stable, and remain intact in the mid gut, and also move across the gut wall into the hemolymph. An alteration in the protein's amino acid content or sequence influences the function of that protein. Likewise, anti-insect activity of a proteolysis- susceptible toxic protein can be improved by administration of protease inhibitors (PIs), which prevent degradation of the toxic proteins, and allows them to exert their defensive function. Better understanding of protein structure and post- translational modifications contributing to stability in the insect gut would assist in predicting toxicity and mechanism of plant resistance proteins. Recent advances in microarray and proteomic approaches have revealed that a wide spectrum of plant resistance protiens is involved in plant defense against insects.
Plant lectins
Lectins are carbohydrate-binding (glyco) proteins, have protective function against a range of pests. The insecticidal activities of different plant lectins have been utilized as naturally occurring insecticides against insect pests. One of the most important properties of lectins is their survival in the digestive system of insects that gives them a strong insecticidal potential. They act as antinutritive and/or toxic substances by binding to membrane glycosyl groups lining the digestive tract, leading to an array of harmful systemic reactions. Lectins are stable over a large range of pH and damage the luminal epithelial membranes, thereby interfere with the nutrient digestion and absorption (Table 1).
Flavonoids
Flavonoids are cytotoxic and interact with different enzymes through complexation. Both flavonoids and isoflavonoids protect the plant against insect pests by influencing the behavior, and growth and development of insects.
Tannins
Tannins have a strong deleterious effect on phytophagous insects and affect the insect growth and development by binding to the proteins, reduce nutrient absorption efficiency, and cause mid gut lesions. Tannins are astringent (mouth puckering) bitter polyphenols and act as feeding deterrents to many insect pests. They precipitate proteins nonspecifically (including the digestive enzymes of insects), by hydrogen bonding or covalent bonding of protein NH2 groups. In addition, tannins also chelate the metal ions, thereby reducing their bioavailability to insects. When ingested, tannins reduce the digestibility of the proteins thereby decrease the nutritive value of plants and plant parts to insects. Role of tannins in plant defense against various stresses and their induction in response to insect damage has been studied in many plants.
Proteinase inhibitors
Proteinase inhibitors (PIs) cover one of the most abundant defensive classes of proteins in plants. Higher concentration of PIs occurs in storage organs such as seeds and tubers, and 1 to 10% of their total proteins comprise of PIs, which inhibit different types of enzymes and play an important role in plant defense against insect. PIs bind to the digestive enzymes in the insect gut and inhibit their activity, thereby reduce protein digestion, resulting in the shortage of amino acids, and slow development and/or starvation of the insects. The defensive function of many PIs against insect pests, directly or by expression in transgenic plants to improve plant resistance against insects has been studied against many lepidopteran and hemipteran insects. The success of transgenic crops in expressing PIs against insect pests has accentuated the need to understand the mechanisms, and interactions of multiple PIs with other defenses, and the adaptive responses of the insects (Table 2).
Enzymes
Enzymes also one of the important aspects of host plant resistance against insects is the disruption of insect's nutrition. The enzymes that impair the nutrient uptake by insects through formation of electrophiles includes peroxidases, polyphenol oxidases, ascorbate peroxidases, and other peroxidases by oxidizing mono or dihydroxyphenols.
Indirect Defenses Methods
The defensive response in plants to attract natural enemies of insects plays a pivotal role in protecting the plants against insect attack. Indirect defenses can be constitutive or induced as a result of combined action of mechanical damage and elicitors from the attacking herbivore. Production of volatiles and the secretion of extra floral nectar mediate interactions of plants with natural enemies of the insect pests (i.e., parasitoids
or predators), which actively reduce the numbers of feeding herbivores. Induced indirect defenses have received increasing attention recently and have been studied on the genetic, biochemical, physiological, and ecological levels.
Herbivore induced plant volatiles (HIPVs)
In this case plants indirectly defend themselves from insect feeding by emitting a blend of volatiles and non-volatile compounds. Insect induced plant volatiles (HIPVs) play an important role in plant defense by either attracting the natural enemies ofthe insects or by acting as feeding and/or oviposition deterrent. HIPVs are the lipophilic compounds with higher vapor pressure which are released from the leaves, flowers, and fruits into the atmosphere, and into the soil from the roots by plants in response insect attack. The HIPV's produced vary according to the plant and insect species, the developmental stage and condition of the plants and the insects. An optimum quantity of volatile compounds is normally released by the plants into the atmosphere, whereas a different blend of volatiles is produced in response to insect. The volatile blend released by plants in response to insect attack is specific for a particular insect-plant system, including natural enemies and the neighboring plants. The HIPVs mediate the interactions between plants and arthropods, microorganisms, undamaged neighboring plants, or intraplant signaling that warns undamaged sites within the plant (Figure 1). Depending upon the modes of feeding of insect pests, different defense signaling pathways are activated, which induce the production of specific volatile compounds [13].
Defense elicitors (insect oral secretion)
Plants undergo a dynamic change in transcriptomes, proteomes, and metabolomes in response to herbivore- induced physical and chemical cues such as insect oral secretions and compounds in the oviposition fluids. It is generally believed that insect-induced plant responses are mediated by oral secretions and regurgitates of the herbivore. The defenses generated by various elicitors differ based on the type of the elicitor and the biological processes involved. A potential elicitor of herbivore-induced plant volatiles from the regurgitate of Pieris brassicae L. larvae has been identified as β-glucosidase which results in emission of a volatile blend from mechanically wounded cabbage leaves that attract the parasitic wasp, Cotesiaglomerata (L.) [14].
Role of phytohormones in induced resistance in plants
Plant defense against insect attack involves many signal transduction pathways that are mediated by a network of phytohormones. Plant hormones play a critical role in regulating plant growth, development, and defense mechanisms. A number of plant hormones have been implicated in intra- and interplant communication in plants damaged by insects. Most of the plant defense responses against insects are activated by signal- transduction pathways mediated by jasmonic acid, salicylic acid, and ethylene. Specific sets of defense related genes are activated by these pathways upon wounding or by insect feeding. These hormones may act individually, synergistically or antagonistically, depending upon the attacker (Figure 2).
Jasmonic acid is the most important phytohormone linked to plant defense against insects and activates the expression of both direct and indirect defenses. Jasmonic acid is derived from linolenic acid through octadecanoid pathway and accumulates upon wounding and herbivory in plant tissues. Chewing of plant parts by insects causes the dioxygenation of linoleic acid and linolenic acid.
Jasmonic acid has also been reported to affect Calcium- Dependent Protein Kinases (CDPK) transcript, and activity in potato plants. CDPKs comprise of a large family of serine/ threonine kinases in plants (34 members in Arabidopsis) and play an important role in plant defense against a variety of biotic and abiotic stresses through signal transduction [15]. In addition to the role played by jasmonic acid in direct resistance against insect pests through the induction of various defensive compounds, its role in indirect resistance has also been well established.
Ethylene is an important phytohormone, which plays an active role in plant defense against many insects. Ethylene signaling pathway plays an important role in induced plant defense against insects and pathogens both directly and indirectly. Ethylene signaling pathway works either synergistically or antagonistically, with jasmonic acid in expression of plant defense responses against pathogens and herbivorous insects. It has been reported that Ethylene and jasmonic acid work together in tomato in proteinase inhibitors expression.
Generally, different plant elicitors induced in plants upon herbivory undergo different signal transduction pathways. For example, Calcium ions (Ca2+), reactive oxygen species, etc.
Conclusion and Future Perspective
In all natural habitats, plants are surrounded by an enormous number of potential enemies (biotic) and various kinds of a biotic environmental stress. Nearly all ecosystems contain a wide variety of bacteria, viruses, fungi, nematodes, mites, insects, mammals and other herbivorous animals, greatly responsible for heavy reduction in crop productivity. By their nature, plants protect themselves directly by developing different morphological structure and by producing some compounds called as secondary metabolites. Plant mechanical defenses act negatively on herbivores insects, diminishing their larval and adult performance. Generally, a plant character may present two or more roles at least in some phase of a plant's life history. I discussed a few cases where pubescence, tissue texture, crystals, latex, waxes and resins are effective against insect herbivores. Secondary metabolites, including terpenes, phenolics and nitrogen (N) and sulphur (S) containing compounds, defend plants against a variety of herbivores and pathogenic microorganisms as well as various kinds of abiotic stresses.
An understanding of induced resistance in plants can be utilized for interpreting the ecological interactions between plants and herbivores and for exploiting in pest management in crops. Since the biochemical pathways that lead to induced resistance are highly conserved among the plants, the elicitors of these pathways could be used as inducers in many crops. The future challenge is to exploit the elicitors of induced defense in plants for pest management, and identify the genes encoding proteins that are up and/or down regulated during plant response to the herbivore attack, which can be deployed for conferring resistance to the herbivores through genetic transformation. However, before using an elicitor effectively in agricultural systems, it is important to understand the chemical changes they induce in the plant, the effect of these chemicals on the herbivores especially in the field, and to see if there is any alteration in plant growth and yield. The Eco- genomic approach which includes association and correlation studies, natural selection mapping, and population genomics enables the estimation of variable selection at loci, and differentiates this from processes acting on the whole genome, such as migration and genetic drift.
From a biotechnological, food-developmental, and breeding point of view, understanding the defense systems of plants and learning how to apply the knowledge is of course of huge interest. For instance, modifications of the jasmonic acid pathway have been proposed [16]. However, due to the extensive crosstalk with other hormone signaling pathways, increased resistance against one certain insect herbivore might result in susceptibility towards another. Furthermore, some defense responses might have negative effects on the environment and humanity as well, as they involve toxic bioactive natural products and proteins reducing digestibility of plant material. Still, reducing the need for synthetic insecticides, by developing crop plants resistant to insect herbivores, would be of significant gain for the food and production industry, both at an economical and environmental level.
To Know More About Trends in Technical and ScientificResearch Please click on:
https://juniperpublishers.com/ttsr/index.php
To Know More About Open Access Journals Please click on:
https://juniperpublishers.com/index.php
0 notes
Photo

Emergence of radicle by rupturing seed coat is called as seed germination. The groeth of radicle and plumule is due to cell extension, division and initiation of several biochemical processes. Rate of respiration increases during germination. It is of two types: Epigeal and Hypogeal germination . . . #agriculture #diversity #plant_world #biodiversity #botany #nature #naturalphotography #india #indianagriculture #harvest #food #study #global #instadaily #agri #agricultureglobal #agriculturefacts #farm #farmer #farmers #agronomy #forestry #plantlover #seedscienceandtechnology #seed #development #agventure007 #insta #icar https://www.instagram.com/p/CEmexYyHcEz/?igshid=1mv2g2kx995b9
#agriculture#diversity#plant_world#biodiversity#botany#nature#naturalphotography#india#indianagriculture#harvest#food#study#global#instadaily#agri#agricultureglobal#agriculturefacts#farm#farmer#farmers#agronomy#forestry#plantlover#seedscienceandtechnology#seed#development#agventure007#insta#icar
0 notes
Text
Assessment of Different Sunflower Genotypes under Agro-Climatic Conditions of District Malakand Khyber-Pakhtunkhwa
Authored by Saif Ullah*
Abstract
The present study was conducted with the aim to evaluate sunflower hybrids under agro climatic conditions of district Malakand. A set of 8 sunflower hybrids was evaluated in randomized complete block design having two replications in spring 2014. Each sunflower genotype was sown in five meter long two rows 75cm apart with P-P distance 25cm. Data were recorded on yield and associated traits such as days to flower initiation, days to flower completion, days to maturity, plant height, head diameter, grain yield and 100 grain weight. Significant differences were observed among the genotypes for all the studied traits. The minimum days (80) to flower initiation were observed in Hybrid-14013 while maximum days (85) to flower initiation were observed in Hybrid-14009. Data regarding flower completion showed that Hybrid-14021 took maximum days (95) in flower completion while minimum days (86) was recorded for Hybrid-14013. Data pertaining to plant maturity compiled that Hybrid 14021 took maximum days (120) followed by one day (119) interval rest of all. A maximum plant height (132cm) was observed in Hybrid-14035 while minimum of 109cm in Hybrid-14013. Data related to head diameter showed that a maximum diameter of 17.80cm was recorded for Hybrid-14021 and minimum of 14.20 cm for Hybrid-14048. Maximum 100 seed weight 5g recorded in Hybrid-14041 while maximum (1666kg ha-1) grain yield were noticed in Hybrid 14021 and minimum yield of (661 kg ha-1) in Hybrid-14048. It is concluded that Hybrid- 14021 performed better in agro-climatic condition of Malakand valley and produced maximum yield.
Keywords: Genotypes; Hybrids; Traits
Introduction
Sunflower (Helianthus annuus L.) is an annual plant native to the Americas. Early settlers grew sunflower for food and garden decoration. European brought sunflower along trade routes to Spain, Italy, Egypt, Afghanistan, China, Russia [1]. It possesses a large flowering head, which is inflorescence of sunflower. The sunflower is named for its huge, fiery blooms, whose shape and image are often used to depict the sun. It has a rough, hairy stem, broad, coarsely toothed, rough leaves and circular heads of flowers. The heads consist of many individual flowers which mature into seeds, often in the hundreds, on a receptacle base.
In Pakistan although it was introduced as an oilseed crop 40 years back but its expansion in acreage and production is fluctuating due to various production and socio-economic constraints. Its seed contains 35-55% oil contents. Research work on this crop has shown that there is great potential of growing it under all the soil and climatic conditions in rain-fed as well as irrigated farming system in different agro-ecological zones (PARC).
The total cultivated area of Pakistan is 20.69 million ha. Out of this cultivated area, 16.48 million ha or 79.65% of the total cultivated area are irrigated. In 1970-71, oilseeds occupied nearly 3% of the total cultivated area, which decreased to 2.5% by 2002-03 [2]. Major share in domestic production of edible oil comes from cottonseed and canola, 67 and 19.6%, respectively. The remaining 13.4% are contributed mainly by sunflower [3]. Edible oil is basic need of every country. In country like Pakistan, About 80% of the total needs are met through imports while only 30% come from local production. Major growing areas of sunflower in Pakistan includes, Multan, Bahawalpur, Sargodha, Faisalabad in Punjab, Peshawar, Malakand in Khyber Pakhtoonkhwa, Khairpur and districts of Hyderabad division in Sindh [4]. During the year 2011-12, the total availability of edible oil was 2.748 million tons. Local production of edible oil remained 0.636 million tons while imports were 2.148 mill tons. The import bill during 2011-12 stood at Rs.216.4 billion (US$ 2.426 billion). During the year 2012-13 (July-March), 1.738 million tons of edible oil valued at PRs. 153.3 billion (US$ 1.595 billion) has been imported. The local production during 2012-13 (July-March) was 0.612 million tons. Total availability of edible oil from all sources is provisionally estimated at 2.35 million tons during 2012-13 (July-March). (Economic survey of Pakistan 2012-13)
Uses of sunflower
I. Sunflower oil is used as edible oil in many parts of the world.
II. Sunflower oil could be used as fuel for cars or trucks.
III. Sunflower oil is an ingredient in salad dressings and cooking.
IV. Sunflowers are used to prepare some special paints.
V. Sunflower oil is used to prepare certain kinds of glue.
VI. Soaps made with sunflower oil.
VII. It has been used in certain paints, varnishes and plastics because of good semidrying properties without color modification associated with oils high in linoleic acid.
VIII. The use of sunflower oil (and other vegetable oils) as a pesticide carrier, and in the production of agrichemicals, surfactants, adhesives, plastics, fabric softeners, lubricants and coatings has been explored
IX. Also use as bird feed.
X. In human diet it is used as snacks and this trend is increased since 15years.
XI. It can be used as a double crop after early harvested small grains or vegetables, an emergency crop, or in areas with a season too short to produce mature corn for silage.
XII. Non-dehulled or partly dehulled sunflower meal has been substituted successfully for soybean meal in isonitrogenous (equal protein) diets for ruminant animals.
XIII. The growing herb is extremely useful for drying damp soils, because of its remarkable ability to absorb quantities of water.
XIV. The Sunflower is a good bee plant, as it furnishes hive bees with large quantities of wax and nectar.
XV. The unexpanded buds boiled and served like Artichokes form a pleasant dish.
XVI. Sunflowers, when the stalks are dry, are as hard as wood and make an excellent fire.
XVII. Of the ash obtained from burning the Sunflower stems and heads (apart from seeds) 62 per cent consists of potash.
XVIII. Being so rich in oil, they are too stimulating to use alone and should only be used in combination with other feeding stuffs.
XIX. The Chinese grow this plant extensively, and it is believed that a large portion of its fiber is mixed with their silks.
Botany of sunflower
The sunflower is a member of the plant family Asteraceae, or Compositae. Asters are remarkable for their type of inflorescence, which is a head of florets. The head is also called capitulum. The “flower” of the sunflower is actually made up of lots and lots of little flowers, called florets. The center, darker florets are disc florets; while the outer, petal-like ones are ray florets. Sunflower seeds are indehiscent achenes. The genus Helianthus, to which the Sunflower belongs, contains about fifty species. It is an annual herb, with a rough, hairy stem, 3 to 12 feet high, broad, coarsely toothed, rough leaves, 3 to 12 inches long and circular heads of flowers, 3 to 6 inches wide in wild specimens and often a foot or more in cultivation. Chromosome number of sunflower is 34. Sunflower is highly cross pollinated crop. For instance, the sunflower genome 3.5 billion bases long [5], slightly longer than the human genome [6].
Flower
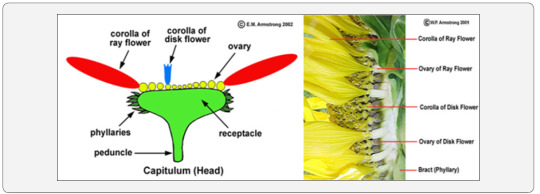
Sunflower is a member of the aster family. A capitulum or head, the characteristic inflorescence of the sunflower family (Asteraceae). The inflorescence consists of ray flowers, disk flowers, or both ray and disk flowers. The ovary of each flower is situated below the attachment of the corolla and stamens, a condition referred to as epigenous or inferior. The disc florets are located in the centre of the composite flower, and the ray florets bear the outer ring of petal-like structures. Ray florets are sterile, and disc florets have both male and female structures, including a single ovary that develops into a sunflower seed. A single flower head may have up to two thousand disc florets, each with the potential to develop into a seed (Figure 1).
Roots system
Sunflowers (Helianthus spp.) have a single taproot and smaller, hairy secondary roots. Sunflower roots usually grow 1 to 3 feet deep, and more than 5 feet long roots are also measured (USDA).
Stem
A sunflower stem sample has many vascular bundles along the edge of its sample. Unlike other plants, fibers are visible next to the vascular bundles. The center part of the sample is called pith, in which most cells are found. Stem have leaves and a terminal head (Figure 2).
Leaves
The leaf of a sunflower is considered a simple leaf, which consists of a single blade. The plumule gives rise to the first leaves of the plant that will go on to grow into organs for transpiration, with the opening and closing of the stomata found within the cell structure of leaves; for photosynthesis, and for other metabolic activities.
Yield contributing traits in sunflower
Seed yield is a quantitative character, which is influenced more from climate and environmental factors in sunflower because of being controlled by large number of genes. To increase seed yield, the study of direct and indirect effects of yield components provides the basis for successful breeding program [7]. Head diameter, 1000 seed weight, plant height are other valuable yield parameters that determine yield improvement in the sunflower [8].
Objective of the study
Present study was carried out to;
I. Check performance of eight sunflower hybrids, obtained from National Agricultural Research Centre Islamabad (NARC) under field conditions of district Malakand.
II. Record data on various quantitative traits.
III. Find out the best sunflower hybrid for the area.
Materials and Methods
A total of 8 sunflower hybrids obtained from NARC oilseed section were sown in the field of district Malakand during February 2014 for yield and associated traits. Plant material was sown in randomized complete block (RCB) design having three replications. Each entry was assigned a two row plot having row to row and plant to plant distance of 75 and 25cm, respectively.
Location
The Research field is located at 34.56 ˚N, 71.96 ˚E, at an altitude of 454m above sea level in the Malakand valley. Malakand is located about 1437 km north of the Indian Ocean, 156km from river Kabul, 96km from river Swat and has semiarid climate. The research farm is irrigated by the Dargai canal from the river Swat. Soil texture is clay loam, low in organic matter (0.87%), extractable phosphorus (6.57mg kg-1), exchangeable potassium (121 mg kg-1), and alkaline (pH 7-8.5) and is calcareous in nature. The climate of the area is semiarid where the mean annual rainfall ranges (400 to 500mm), 60-80% rainfall occurs in summer, while the remaining 30-40% rainfall occurs in winter (Table 1).
A. Parameters studied: Data were recorded on the following parameters.
B. Days to flowering initiation (DFI): Days to flowering initiation was recorded from date of sowing till about five percent of the buds opened flowers in each plot.
C. Days to flower completion (DFC): Flower completion data was recorded when about 90% of the buds opened flower in each plot. This was also calculated from date of planting.
D. Days to Maturity: Data for days to maturity was recorded when back of the heads turned yellow and bracts started turning brownish in color.
E. Plant height (cm): A total of five plants randomly selected in each plot were measured from ground level to attachment of head with stem to record data on plant height. Plant height was recorded at the time of maturity.
F. Head diameter (cm): Head diameter was calculated as it is yield contributing trait in sunflower. Five plants from each line were selected at random and data for head diameter was recorded. Head diameter was measured from one edge of the disk to other.
G. 100-seed weight (g): 100 seed weight was also calculated for each hybrid. An average of three samples of 100-seeds for each hybrid, were taken to record data on 100- seed weight. Seed was weighed in grams with the help of an electronic balance up to two decimal points.
H. Grain Yield (kg ha-1): Total produce after threshing was sun dried for 2-3 days and then seed was weighed with the help of an electronic balance for each hybrid.
I. Statistical analysis: Data after compilation was statistically analysed using STATISTICA 8.1 software and means were separated by LSD test.
Results and Discussion
Days to flower initiation
Analysis of variance revealed highly significant (P=0.01) differences among all the Genotypes (Table 2). Days to flower initiation ranged from 80-85 days. Hybrid-14001 took minimum days (80) to flower initiation whereas Hybrid-14009 took maximum days (85) for flower initiation (Table 3). These results are partially in line with those of Shah [9]. The differences in results may be due to differences in Genotypes or due to environmental effects.
Days to flower completion
Analysis of variance revealed that days to flower completion showed significant (P<0.05) differences among all the Genotypes (Table 2). Days to flower completion ranged from 80-92 days. Hybrid-14023 took minimum days (86) for their flower completion while Hybrid-14035 took maximum days (92) to complete their flowers (Table 3). Furrkh [4] also found significant variation for days to flower completion.
***Significant at 5 and 1% level of probability respectively, whereas NS= Non-significant.
Days to maturity
Analysis of variance revealed that days to maturity showed non-significant (P>0.05) differences among all the Genotypes (Table 2). Days to maturity ranged from 119-120. Hybrid-14005 took minimum days (119) to be matured whereas Hybrid-14021 took maximum days (120) to be matured (Table 3). Khalid [8] also found non-significant differences for days to maturity.
Plant height (cm)
Analysis of variance revealed that plant height showed nonsignificant (P>0.05) differences among all the genotypes (Table 2). Plant height ranged from 110-135cm. Hybrid-14013 had minimum (110cm) plant height whereas Hybrid-14021 had maximum (135cm) plant height (Table 3). Khalid [8] found the plant height to be non-significant.
Head diameter (cm)
Analysis of variance revealed that head diameter showed significant (P<0.05) differences among all the Genotypes. Head diameter ranged from 14-18cm. Hybrid-14048 had minimum (14cm) head diameter while Hybrid-14021 had maximum (18cm) head diameter. These results confirmed the earlier findings of Arshad [1] also found significant variation for head diameter.
Grain yield (kg ha -1)
Analysis of variance showed non-significant (P>0.05) differences for grain yield among all the genotypes (Table 2). Grain yield ranged from 817-1666kg ha-1. Hybrid-14041 had minimum grain yield (817 kg ha-1) whereas Hybrid-14021 had maximum grain yield (1666 kg ha-1) (Table 3) Khalid [8] also found Non-significant relation for Grain Yield kg ha-1.
100-grain weight (g)
Analysis of variance revealed that 100-grain weight showed non-significant (P>0.05) differences among all the genotypes (Table 2). 100-grain weight ranged from 04-05g. Hybrid-14013 had minimum 100-grain weight (04g) whereas Hybrid-14041 had maximum 100-grain weight (05g) (Table 2). Anjum [1] also got non-significant difference for 100 grain weight.
Conclusion and Recommendations
There is sufficient genetic variability in the tested sunflower hybrids which is useful tool for grouping of different hybrids according to their adaptation through the environmental condition, Hybrid 14021 produced highest grain yield (1666kg ha-1) followed by hybrid 14009 (1517kg ha-1) and thus are considered best among all hybrids evaluated during this study. From the above study and data obtained from results it is recommended that Hybrid 14021 and 14009 fits well in the agroclimatic conditions of Malakand valley and produce maximum yield [10-12].
To Know More About Current Trends in Biomedical Engineering & Biosciences Please Click on: https://juniperpublishers.com/ctbeb/index.php
To Know More About Open Access Journals Publishers Please Click on: Juniper Publishers
0 notes
Link
Salinity in soil or water is one of the major stresses that affect crop production around the world. In Bangladesh the coastal areas are increasing day by day due to climate change. Therefore it is very important to investigate the mechanisms of salt tolerance. That is why, this study was undertaken to investigate the effect of salinity on tomato (Lycopersicon esculentum Mill.) by using ten genetically diverged tomato genotypes during seed germination and seedling growth stage. The study was carried out in Completely Randomized Design (CRD) with three replications under invitro condition. In the study, emergence percentage, radicle length, plumule length, Proline content, K+/Na+ of the seedling were assayed on five levels salinity; control (0), 4,8,12 and 16 dS m-1
0 notes
Text
Mobile phone radiation overwhelms the antioxidant systems of mung bean root cells.
PMID: Sci Total Environ. 2009 Oct 15 ;407(21):5543-7. Epub 2009 Aug 13. PMID: 19682728 Abstract Title: Mobile phone radiation inhibits Vigna radiata (mung bean) root growth by inducing oxidative stress. Abstract: During the last couple of decades, there has been a tremendous increase in the use of cell phones. It has significantly added to the rapidly increasing EMF smog, an unprecedented type of pollution consisting of radiation in the environment, thereby prompting the scientists to study the effects on humans. However, not many studies have been conducted to explore the effects of cell phone EMFr on growth and biochemical changes in plants. We investigated whether EMFr from cell phones inhibit growth of Vigna radiata (mung bean) through induction of conventional stress responses. Effects of cell phone EMFr (power density: 8.55 microW cm(-2); 900 MHz band width; for 1/2, 1, 2, and 4 h) were determined by measuring the generation of reactive oxygen species (ROS) in terms of malondialdehyde and hydrogen peroxide (H(2)O(2)) content, root oxidizability and changes in levels of antioxidant enzymes. Our results showed that cell phone EMFr significantly inhibited the germination (at>or =2 h), and radicle and plumule growths (>or =1 h) in mung bean in a time-dependent manner. Further, cell phone EMFr enhanced MDA content (indicating lipid peroxidation), and increased H(2)O(2) accumulation and root oxidizability in mung bean roots, thereby inducing oxidative stress and cellular damage. In response to EMFr, there was a significant upregulation in the activities of scavenging enzymes, such as superoxide dismutases, ascorbate peroxidases, guaiacol peroxidases, catalases and glutathione reductases, in mung bean roots. The study concluded that cell phone EMFr inhibit root growth of mung bean by inducing ROS-generated oxidative stress despite increased activities of antioxidant enzymes.
read more
0 notes
Text
Global Luxury Duvet Market 2019-2024
Global Luxury Duvet Market 2019-2024
Down Duvets are made from the light, fluffy clusters and plumules that come from beneath the feathers of ducks and geese. The quality of down can be determined by the fill power. Down Duvets are great options for anyone allergic to down feathers or for anyone on a stricter budget.
According to this study, over the next five years the Luxury Duvet market will register a xx% CAGR in terms of…
View On WordPress
#Luxury Duvet market#Luxury Duvet market analysis#Luxury Duvet market forecast#Luxury Duvet market growth#Luxury Duvet market overview#Luxury Duvet market research#Luxury Duvet market share#Luxury Duvet market size#Luxury Duvet market study
0 notes
Text
It's here in Uncle Nekk's eyes too. A sadness-sympathy-pity Kido can't seem to escape from no matter where he turns, not even when he's with the other kids, or when he's tucked himself away in his aerie, or when he's here, right now, trying to move on from— from someone who clearly moved on from him a long time ago.
It shouldn't make him this blisteringly angry. Like there's some sapsucker nipping away at a point in his heart he's just fixed: a split leaking something hot and bubbling and sticky with bitterness.
The wood clutched between his feathers doesn't even creak, but Kido likes to imagine, for a heartbeat, that it splinters into irreparable pieces.
"How much," he says, breaths as measured as he's practiced in the dark of an empty aerie, "can I get for this?"
Just like Auntie Misa when he tried this with her, Uncle Nekk looks away for a moment. "Kido, I don't think—"
"How much," he grits out, and it's not very polite, it's not very nice, but Kido hasn't felt very polite or very nice for a while.
Uncle Nekk stops looking away. He's frowning, but in the way all the grown-ups are trying not to when they're with Kido — not-frowning so hard it circles right back to it. He doesn't say anything for a few seconds, just sighs and shakes his head, before he's coming around the checkout desk.
"I don't think," Uncle Nekk starts, kneeling to level their gazes and place a wing on Kido's shoulder, "you want to do this."
A really, really scary thought flits through Kido's mind, and he's backing out of that gentle touch with a shriek clogged in his throat.
Of course. Of course. Kido's the only one who feels how deep this hurt really goes, how bad this fear-anger-grief can get, how much he wants to do this — to get rid of this painful reminder in a way that will at least get him something back from the world-upending loss — and it's Uncle Nekk who knows better. Like how it was Auntie Misa who knew better, and Uncle Gesane who knew better, and probably every other adult in this flock who knows better. Of course.
"How MUCH," he tries again, voice bolstered by the wave of barely contained resentment washing over him, and it's so, so freeing and so, so terrible.
"Oh, Kido," Uncle Nekk says — like Auntie Misa and Uncle Gesane and every other adult in this flock — soft and quiet and not at all annoyed.
It's. It's so much, it's too much, it's what made Kido flee from Auntie Misa in the first place, except he can't even do that here 'cause Uncle Nekk's got both his shoulders now. He can feel the oncoming burn of stupid tears when they're the very last things he needs or wants right now — he's not a hatchling anymore, he's not, but he's done so much stupid crying since— since that he might as well be one again, and the more he lets his tears say the things he won't, the more the grown-ups will treat him like he's incapable of even knowing what he wants, and— and—
"Can you just—!" Kido stutters, shoving his— the Feathered Spear in his grasp out like he can't stand to hold it any closer, "I don't want this! I don't— I can't have it, I don't wanna have it!"
The tears fall, as they always do, and Uncle Nekk says nothing. The silence stretches, reaching for the rest of Kido's words, and he's helpless when it plucks them all out between gasps and wheezes, "Please, I just— wan-na stop thi-inking about it and— and rem-remembering and— I don't— I don't! Want it! I don't want it!"
Somewhere in the middle of Kido making a complete embarrassment of himself, Uncle Nekk takes the spear from him. Somewhere in the middle of Kido crumbling for the last time — the last time, please — Uncle Nekk pulls him into his wings in a maybe-sorta-kinda-hug.
"Okay," he says, hesitantly, like Tulin does when he doesn't know what else to say. It's funny how his voice wobbles — or it would be, if Kido wasn't so distressed. "How...how about this. How about I hold onto this for you for now, huh? Until you decide you want it back."
He won't want it back, ever, and he opens his beak to say exactly that — but Uncle Nekk beats him to it.
"Which can be never! That...works, too."
Good. Good. Some tiny part of him even thinks, it better.
In the quiet that follows, the maybe-sorta-kinda-hug dips from bearable to awkward. Kido makes to pull away and Uncle Nekk lets him.
"Thanks," he murmurs, swiping remnants of his tears away, just downcast now that he's gotten what he's come for. He pulls further, intent on shuffling back to his aerie to get himself together again, but— Uncle Nekk? Doesn't let go of his shoulders?
"Kido," Uncle Nekk says, and it's like Cree when she's figured something out now. Firm and determined, though for what, he can't tell. "Why don't you stay a while? I could use an extra wing right about now — and, you know, I've missed your little visits."
He carefully does not reply. He doesn't think anybody's missed him with how he's been, lately.
"I mean it! Come on, help your Uncle Nekk out? Please?"
Well...when he's being asked so nicely...how can he say no?
#* plumule / study.#( there's! a lot of context here that i may or not may not explain if anyone's interested slkfjkld BUT THIS IS PROBABLY HOW KIDO'S#APPRENTICESHIP WITH NEKK STARTED...#< i say this like i'm not in complete control of what happens to this kid slkfjdlk#the general gist is kido is Hurting and nekk sees that and wants to help out even if only by providing an outlet for all the complicated#emotions kido's clearly feeling but not saying (through the complex-turned-monotonous motions of handiwork)#ho i am almost afraid of posting this....BUT HERE WE GO SEND POST#the only. ONLY thing i may srb later slkfjkdlf )#* reprise / ic.#* ic / para.#long post cw#* v / song of the reclamation.
11 notes
·
View notes
Text
this will only make sense to me but there was probably a period in kido’s life where hearing apologies made him feel worse than anything. something something it was so often said to him by adults who wanted to make things better for him but couldn’t really do anything to “make up for” the absence of his dad and he got so sick of having to assure them that it was fine. it was alright. it’s okay. instead of exploding about it that he developed a Bit of an aversion to hearing sorry from any adult
#* plumule / study.#* roosting / ooc.#his intense faith in his dad returning VS his exhaustion at being the only one who has this faith: fight fight fight!!!#i’ll revisit this later because i literally only just woke from dead sleep skdkfkf#but hi good morn! hopefully will be feeling way better in a few hours since I’ll be getting aches massaged out ahaha#hope to get some writing done today!! have a lovely day/night folks; be seeing you <3
8 notes
·
View notes
Text
wow. finally here to talk about my take on forms of rito languages!! it's a doozy, folks. there will be four i'll be exploring: rito script, rito shorthand, rito key, and birdsong. all under the cut, so here we go!
RITO SCRIPT
i believe this would most likely be cuneiform in nature, if only due to the constraints of rito morphology. i doubt they have any innate, very fine motor control in their wings ( "hands" ) given how big their feathers ( "fingers"? ) are, so impressions and/or inked signs that can be made with wedge-tipped styluses seem a little more logical than the more motion-heavy strokes needed for, say, something like hylian script — which they still use around the village for the sake of non-rito visitors ( notably carved into wood ), but many rito are better readers than they are writers of hylian script! not all, but that's the general trend!
okay, now...i am not a linguist. but i'm thinking the old system ( like. prior to, during, and a little after dineli's time ) was more logographic, and was used way, WAY less than rito was spoken. over the millennia, through language contact ( especially applicable to the rito, who were traditionally nomads and a bulk of whom are still travellers by nature ) and the natural evolution of their spoken analogue, rito script has developed into more of a logosyllabary! still used notably less than its verbal equivalent, but not to the extent of back then; all fledglings in rito village are taught to a decent level of literacy now.
i don't have a physical demonstration in mind because i don't have the brain juices for creating a whole conlang...but i want to emphasise that this script is reflective of spoken rito, which is a tonal language ( THIS IS IMPORTANT ) and not alphabetical.
RITO SHORTHAND
rito stenography of rito script! finds even rarer cases of use than its longhand, but that's just how it is with most shorthands, isn't it? penn is especially proficient in this — his draft notes for reporting are exclusively in this shorthand. makes it harder for those who might want information when he isn't willing to give it to read, and speed is a critical thing when you're on the field! also his notebook is tiny. his pen when it's in his "hand"? tiny. he is not writing longhand in that thing.
i have a more concrete image in my head about this system! it's primarily informed by and based on modern musical notation!! it would still vaguely resemble rito script ofc, but i think this would be neat given how music-entrenched rito culture is...and it's fun to think about possible uses for it as a code! is it sheet music or a message? it will be obvious to rito ( even to those unfamiliar with shorthand, if only in that "huh, this does not read like music" sense ), but to most non-rito? a mystery they don't even realise is a mystery slkfjkdf
RITO KEY
separate from any scripts; this is nonverbal rito communication! misleading title, i know ( <- weak for connecting music to the birdies in any way i can ). their signed language, so to speak, in that words and meanings are conveyed through manual/physical articulation rather than verbal — but there are still some vocal aspects to it as well? very little, and less defined in that it's just like...whistles, trills, warbles, and other actual bird noises ( almost dipping into birdsong ). these aren't necessary most of the time; they're used to accentuate or clarify signs, not outright replace them.
i like to think that their signs, when grounded, place heavy emphasis on wings, shoulders, head positions, etc. there's not a lot of individually bending wingtips/"fingers" — "hand" movements are mostly ( but not all ) fully splayed or fully clenched, if that makes sense? and facial expressions would be very important too, as it is with most signed languages.
and signs when flying...i'm thinking maybe flight patterns? possibly whole flight motions? this is where the vocal side of rito key might really come into play, but again isn't strictly necessary — especially if you're a good flier. does convey less specifics than its grounded form though.
most grown travellers and warriors know rito key. it's a useful language to learn! can offer the advantage of subtle but active coordination on the battlefield and such.
BIRDSONG
this is, essentially, "birdspeak". natural language of birds, including rito! penn proved that rito can speak to birds to such a depth that they are his primary informants, and even though literally nothing in the game indicates this is a universal thing, i am RUNNING with this tidbit and saying yeah. this is an inherent thing for the rito. not in that they're born with an automatic understanding of it, but in that it's the easiest thing for them to learn. like their first language! do you see my vision? do you see it! i get to decide this, it's my sleepover!! ( affectionate )
this is much more simplistic than any other language a rito would know; there are no specific words that would equate to specific meaning. it's all ( bird ) sounds! no nuance to this language. it's very direct, straightforward, and in-the-moment. a major case of interpretation =/= translation, which is why an actual rito language naturally arose sdlfsjdfk
#* arrowhead / study.#* plumule / study.#* powder down / study.#* field notes / study.#* wisps / study.#* roosting / ooc.#not me scouring this thing for the 10th time for anything that could be a contradiction in my thoughts or previous studies lsfkdlg#i think. we are good. i think we are good.#I AM HITTING POST. HERE WE GO. WOE WALL OF TEXT BE UPON YE#ngl though i had fun compiling my thoughts this time :)#special thank you to hilary for getting me to think about rit.o key!!#long post cw#* vanes / study.
12 notes
·
View notes
Text
everybody's asleep! time to post kido thoughts because i have so many things brewing in my head about him that i can never seem to word to my satisfaction and i am finally taking the leap and treating him like my canon muses in that you know what? the studies focusing solely on him do not have to be neat either!!
loves his dad. hates his dad. loves his dad enough to keep the quill from his feathered spear — the last physical connection to him he's got — to bind to the braided cord he receives from nekk for his growing semiplumes as a statement of you are still something to me. hates his dad enough that he never gave his weapon its proper rites by burning it in a funerary bonfire, which wouldn't be a problem if he also did not refuse to take it up in any shape or form ( nekk's still got it in his aerie, collecting dust ). it's complicated.
has healed enough from the loss that he can think of his dad and anything to do with him without ever feeling like his stomach's being pried open to be turned inside-out by the time totk rolls around. still wounded to the point of rejecting customary warrior training and finding that extensive, emphasised talk of warriorhood in general, a literal cultural hallmark, makes him genuinely anxious and even a little angry. just a little. he can at least immerse himself in the mythic touch and triumph of Songs.
loves revail. hates revali. still has all his Songs memorised and could recite the older ones by heart, still gets dreams of being buoyed up above the skies by what he imagines would be his gale, still quietly thinks he's one of the coolest people peaks among the ripples will ever have to grace their history. also can't feel normal about him in any single conversation. also gets irrationally mad when anyone goes on and on about him, the ( mostly silent ) critic to the enjoyer. also rather unhealthily, if only subconsciously, clings onto the idea that his dad chose revali and his legacy over himself — not in the "oh, i was not enough" way ( though he did have a stint of this ), but in the "oh, of course he'd choose someone so amazing like master revali over me" way. the idea of revali is the last thing connecting him to his past with his dad, where he was still present, and kido hasn't decided whether he wants to burn that bridge or reinforce it.
but hey, here's to fluffier things! is contender for the one with the best fine motor control among his peers. though he doesn't feel a passion for it, and would much rather something else at his current age, he does hold his clothier apprenticeship with nekk near and dear to his heart for giving him an outlet he very sorely needed and the inability to ( physically and emotionally ) self-isolate as he wanted to in the worst of his grief.
cartography came a bit out of nowhere, but at the same time it's perfectly fitting. he legitimately enjoys travelling, even if his initial taste for it came in little bites of attempted-and-failed rescue missions. and maybe he wants to make sure he'll be able to find anyone he cares about in the future, huh? maybe he wants to be able to pinpoint exactly where anyone could've gone no matter what. bonus points: it's an artisan's life, not a warrior's one! score! ( < is going to be so upset when he finds out that learning to at least know how to defend himself is a compulsory step in graduating from fledgling to globetrotter )
#* plumule / study.#* roosting / ooc.#some of these are part of his innermost thoughts and feelings that you will not be able to pry out of him unless you're looking for them#so like. not even applicable to threads. but you know me and meta that most likely won't come up in threads!#i realise the vibes with kido are. very somber. but i promise you he's a more joyful kid than my recent studies make him out to be slkfjkld#i am experiencing the horrors of executive dysfunction and so am in a bit of a Mood but!! i hope to be back for dms and writing!#hope you're having a lovely day folks! be seeing you <3#child neglect implied
9 notes
·
View notes
Text

well. we done did it! thank you @gloryseized for volunteering your link ( you didn't even need to say anything. i just took your like and ran with it <3 ), muses' thoughts under the cut :)

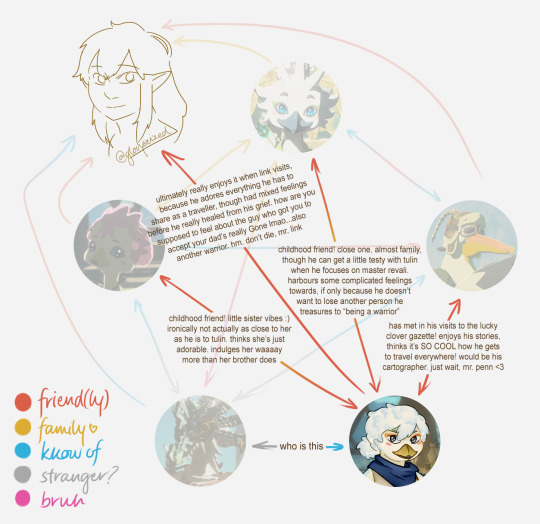
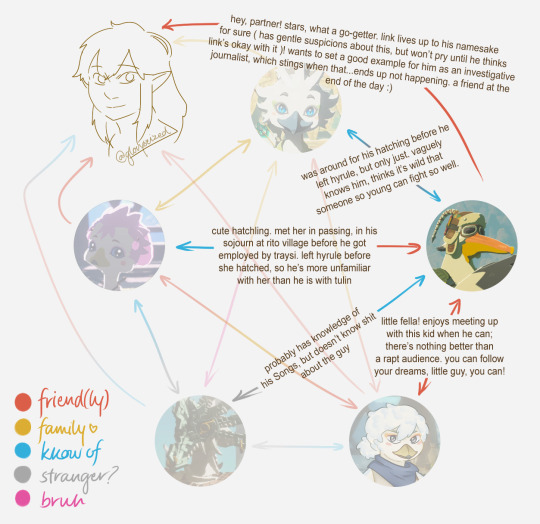
#* arrowhead / study.#* plumule / study.#* powder down / study.#* field notes / study.#* wisps / study.#* roosting / ooc.#i did have lots of fun with this!! even if it's only anybody -> li.nk's thoughts that actually matter in the long run skfjldkf#this actually consumed my time so bad so i must bid y'all goodnight :'D more writing...another time!#have a wonderful rest of your day folks! be seeing you!#long post cw
9 notes
·
View notes
Text
i'm sure i've made a few insinuations about this already, but it's probably high time i finally talk about my take on it; plumage ornamentation is typically a form of allopreening in any given rito flock, and hence holds lots of social and cultural importance for them!
(which isn't to say that it's always allopreening or can't be autopreening — some ornamentations are just part of rito tradition and denote a role or status — but a rito who isn't allopreened even semi-regularly can uh. be a very lonely rito. it can get pretty complex honestly, so it depends on what's actually going on!)
in addition to the mechanics of their own language, ornamentation is another way of casually enforcing bonds and dynamics for the rito. it's on average most often engaged in between family, but not exclusively reserved for them! in trusting and being willing to let someone touch your feathers and/or being allowed to touch someone else's feathers so intimately, you're letting them (and everyone else) know that they're some form of Really Important to you! kind of like "marking" them? it's at once less profound than i'm making it sound yet still extremely, extremely significant sklfjkf
when communicated through direct physical contact, ornamentation is primarily concentrated on crest feathers and semiplumes, which can translate to hair, fins, or simply the head for non-rito. it involves acts like braiding, beading, binding, and general weaving-things-into-feathers. while the act itself implies a level of comfortable, casual intimacy, what's actually used as decoration can give another layer of meaning to it too!
gifting items and materials for ornamentation holds less intimate weight than actually physically handling someone's feathers, but gets the same interpersonal message across! which is how travelling rito, especially those who go at it solo, are able to keep their loved ones close and basically auto-enforce those bonds with them :)
when interpreting plumage ornamentation (in this context), it's good to keep in mind that:
colours of cords, ribbons, beads, etc. are always chosen very deliberately by the older folks. children are encouraged to experiment more, to get a feel for what they want to express (and for what looks good).
level of skill does not always correspond to level of care (younglings are notorious for being very messy with their affection), but general rule of thumb is that the more eye-catching the work, the more you (and everyone else) are being told this person is Cared For!
moulted rito quills are, for a lack of better words, blessings. specific meaning depends on what sort of relationship the receiver has with the rito whose quill is being gifted, or with the rito who's doing the gifting if they're not the same person, but there are always hints of i'm with you always.
the meanings of ordinary bird quills are defined by their colour!
sometimes, things like flowers, plants, and other non-permanent materials will be used just for fun! doesn't lessen the significance of what allopreening can mean :)
ESSENTIALLY, if a rito asks to do something with your feathers (or whatever is equivalent), they are asking if it's okay for them to let the world know you are Cherished. and sometimes — if you're close enough — they don't even ask, they'll just do it!
#* arrowhead / study.#* plumule / study.#* powder down / study.#* field notes / study.#* roosting / ooc.#(re: the last line — tuli can be an absolute fiend about that slkjfd)#god i hope this all makes sense sdlfkjdld using 'preening' more loosely than its literal definition#but finally fought through all my articulation issues to give my two cents on this!#AND THIS IS JUST MY THOUGHTS ON PREENING IN THIS CONTEXT....some things here are applicable to ornamentation in a non-interpersonal sense#and i have things lurking in my head about materials like arrowheads and gemstones and other carved and cut headpieces and!!#i still want to work out a colour-meaning system for them.....given how strong their photopic vision is#i don't see why there wouldn't be an emphasis on colour (at least with these things) in addition to the oral/musical aspect of their cultur#WHICH I ALSO...NEED TO MAKE A POST ON....Songs i will get to you i prommy lksjdfkl#long post cw#* wisps / study.#* vanes / study.
9 notes
·
View notes
Last Seen Blogs

portariaremotasopaulo508
portaria remota São Paulo

retquits
we met once long ago

spookyscylla
the sassy sea beast ™

keggsofsanmiguel
keggplant

cnradz
baguette