#because of how significant it is to the history of palaeontology
Text
Ok y’know what I haven’t made a reblog game before and I wanna know so,,
Reblog and put in the tags what your favourite prehistoric animal is and why!
#dinosaurs#paleontology#palaeoblr#reblog game#mine is sinosauropteryx#because of how significant it is to the history of palaeontology#not only the first non-avian dinosaur described with feathers#but also one of the first and best fossils that preserves colour!!#they looked like little birdy red pandas and I love them so much#they even have a bandit mask like a raccoon!!#blessed creature
206 notes
·
View notes
Text
Evolutionary origins of the pygmy right whale
Researchers have finally settled a decades-long dispute about the evolutionary origins of the pygmy right whale.
The smallest of the living baleen whales, it’s tank-like skeleton is unique, and its ecology and behaviour remain virtually unknown.
Because it is so unusual, the evolutionary relationships of the pygmy right whale (Caperea marginata) have long been a bone of contention.
In a study that solves the debate, just published in Marine Mammal Science, an international group of researchers sequenced the complete genome of Caperea, combining their findings with morphology and palaeontology.
Co-author Dr Felix Marx, curator of marine mammals at the Museum of New Zealand Te Papa Tongarewa explains the skull shape of the pygmy right whale seems to be adapted for skim-feeding, where a whale will swim at the water’s surface with its mouth open to food.
“This is very similar to the larger true right whale, leading some scientists to believe the two whales are closely related, hence their similar names. However, others believe the pygmy right whale is more closely related to species like the blue whale, which take big gulps of water to collect food instead of skimming,” he says.
He was pleased to be able to exploit the power of genomics to elucidate the history of life.
“After 150 years of anatomical orthodoxy and decades of dispute, genomics now shows beyond reasonable doubt that Caperea is a distinct lineage and not related to right whales.
“Like river dolphins and sperm whales, Caperea is the sole guardian of a unique piece of evolutionary heritage. It’s not just another weird right whale — it truly is the last survivor of an otherwise lost family that once played a much bigger role in Earth’s history,” he says.
Co-lead author Dr Kieren Mitchell, of Manaaki Whenua — Landcare Research, says new genetic information often prompts scientists to reconsider why different animals appear more similar or different to each other.
“When DNA and anatomy seem to be at odds about the relationship between species, usually that means there’s an even deeper and more interesting story to be discovered about their evolution,” he says.
Co-author Dr Nic Rawlence, Otago Palaeogenetics Laboratory Director, describes Caperea as a “wonderful case of convergent evolution,” which occurs when two unrelated species end up appearing more and more alike as they adapt to similar selective pressures.
“Caperea has historically been aligned with right whales because they look the same due to similar feeding strategies, when, in fact, it’s probable that Caperea is the last surviving member of an ancient group of whales called cetotheres,” he says.
Co-lead author Dr Ludo Dutoit, of Otago’s Department of Zoology, says now its position in the family tree of whales has been confirmed, researchers can start to explore what the Caperea lineage looks like, and what kind of past events were significant in driving its evolution.
Dr Marx agrees, adding that Caperea may be another example of how being ‘unusual’ helped save a lineage from extinction.
“River dolphins likely survived the demise of their marine relatives because they invaded freshwater habitats; sperm whales persisted when their toothed relatives disappeared because they were deep-diving suction specialists; and Caperea survived because it adapted to be a skim filter feeder, when most of its relatives presumably didn’t.”
0 notes
Text
Folded Forest: Defining the Jurassic Period
by Jane Thaler
What’s in a Name?
Derived from the words for “middle life” in Greek, the Mesozoic Era consisted of three geological periods: the Triassic, Jurassic, and Cretaceous. While many of us might be aware of all three, the term “Jurassic” has seeped into our everyday lives in a way that the Triassic and Cretaceous have not. We can attribute much of this ubiquity to the wildly popular Jurassic Park books by Michael Crichton and their subsequent film adaptations, but have you ever wondered what “Jurassic” actually means and how scientists define the period’s geological boundaries?
Coining “Jurassic”
The “Jura” in Jurassic refers to the Jura Mountains that run along a large portion of the Swiss and French border. Named for the ancient Celtic word for forest, the Jura Mountains are known for their tree-covered peaks and the folded rocks that comprise them (Jones, 2020, p. 94).

Jura Mountains from Wikimedia Commons.
It was here in 1795 that Alexander von Humboldt, a Prussian explorer and naturalist, documented a series of carbonate shelf deposits from the period now known as the Jurassic and dubbed them the “Jura Kalstein.” Alexander Brongniart, a French scientist known for arranging and describing the geologic formations of the Tertiary Period (66.0 to 2.6 million years ago), coined the term “Terrains Jurassiques” to refer to all Jurassic strata in 1829. In 1832, German geologist Leopold von Buch established the three-fold subdivision of epochs based on the folds of limestone in the Jura: the Lias (Early Jurassic), the Dogger (Middle Jurassic), and the Malm (Late Jurassic). This arrangement remains the basic framework for our geological understanding of the Jurassic to this day (Ogg et al., 2012b, p. 732; Encyclopedia Britannica, 2021a).

Jura Mountain fold known as the “Chapeau de Gendarme” from Wikimedia Commons.
Beginning of the Jurassic
Nestled between the Triassic and Cretaceous periods, the Jurassic spanned from 201.3 million years ago to 145 million years ago (National Park Service, 2020). The end of the Triassic (so named because it is a group of three strata) and the beginning of the Jurassic is marked by the Triassic–Jurassic (Tr–J) extinction event, sometimes called the end-Triassic extinction. The fourth of five major extinction episodes on Earth (or sixth if you count the current, anthropogenic extinction), the Tr–J extinction wiped out around 75 percent of all marine and terrestrial life (Encyclopedia Britannica, 2021b).
Current evidence suggests that the Tr–J extinction was initially set into motion by movements of the Earth’s crust. As the all-encompassing mega-continent Pangea began to break apart, the associated tectonic shifts caused significant volcanic activity that spewed carbon dioxide into the atmosphere. The resulting global warming disrupted the Earth’s carbon cycle and contributed to ocean acidification (Fuge, 2020).
The ecological niches left open by the Tr–J extinction were quickly filled by remaining species of pterosaurs, crocodilians, turtles, mammals, many species of plants and invertebrates, marine life, and dinosaurs. Though many species died out during the extinction event, the wet and warm climate of the Jurassic in many places encouraged the growth of lush vegetation along with the proliferation and diversification of fauna. Oceans teemed with life, forests flourished, and dinosaurs became the dominant forms of backboned animal life on land during this time (Encyclopedia Britannica, 2021b).

Jurassic landscape in the Dinosaurs in Their Time exhibition at Carnegie Museum of Natural History.
End of the Jurassic
The end of the Jurassic is a bit of a mystery as the geological boundary between it and the Cretaceous Period (the latter name derived from the Latin for “chalk”) remains formally undefined. In fact, the Cretaceous is the only period in the Phanerozoic Eon (541 million years ago to present day) that “does not yet have an accepted global boundary definition” (Ogg et al., 2012a, p. 795). This definitional challenge is due to a number of factors but is mostly attributed to the concept of provincialism or provinciality, which means that plant or animal populations were restricted to a particular area or group of areas (Gale et al., 2020). This resulted in endemic populations, particularly of ammonites, which left uneven or unclear fossil markers in the stratigraphic record (Wimbledon, 2017; see Énay, 2019 for more detail on the J/K boundary debates).
We do know that the end of the Jurassic was marked by the Tithonian–early Barremian cool interval, which began 150 million years ago and continued well into the Early Cretaceous (Ogg et al., 2012a). During this time, some groups of animals did go extinct or become less diverse, like the dinosaurian subgroup Stegosauria that included Stegosaurus, while others increased in abundance, like some ammonite subgroups who survived the Tr–J event. Plants were also developing in important ways during this time. Around 130 million years ago, angiosperms (flowering plants) began to diversify, and they became increasingly dominant throughout the Cretaceous (Friis et al., 2010). Taking the unknowns and variables into account, the end of the Jurassic is currently placed at 145 million years ago.
Naming Geological Periods
Many of the names we still use for geological periods went through a similar process to that of the Jurassic: a scientist named a phenomenon based on the strata they were studying and the nomenclature (the system of names) developed from there. Nowadays, defining and naming geological units is left to the International Commission on Stratigraphy of the International Union of Geological Sciences. The process by which this happens feels about as long as the geologic periods themselves, at least to those of us watching from outside the commission. This is, of course, an exaggeration, but it does take years of work and rounds of voting to arrive at an official stratigraphic boundary designation.
Check out https://stratigraphy.org for the latest updates on humanity’s understanding of geologic time.
Jane Thaler is a Gallery Experience Presenter and Floor Captain in CMNH’s LifeLong Learning Department. Museum staff, volunteers, and interns are encouraged to blog about their unique experiences and knowledge gained from working at the museum.
References:
enay, Raymond (2019). The Jurassic/Cretaceous System Boundary is at an impasse: Why not go back to Oppel’s 1865 original and historic definition of the Tithonian? Cretaceous Research. https://doi.org/10.1016/j.cretres.2019.104241.
Encyclopedia Britannica (2021a). Alexandre Brongniart. https://www.britannica.com/biography/Alexandre-Brongniart
Encyclopedia Britannica (2021b). End-Triassic extinction. https://academic-eb-com.pitt.idm.oclc.org/levels/collegiate/article/end-Triassic-extinction/474417
Fuge, L. (2020). Volcano link to end of Triassic extinction. Cosmos. https://cosmosmagazine.com/history/palaeontology/volcano-link-to-end-of-triassic-extinction/
Friis E. M., Pedersen K. R., Crane P. R. (2010). Diversity in obscurity: fossil flowers and the early history of angiosperms. Philosophical Transactions of the Royal Society B, 365. https://doi.org/10.1098/rstb.2009.0227
Gale, A. S., Mutterlose, J., Batenburg, S., Gradstein, F. M., Agterberg, F. P., Ogg, J. G., Petrizzo, M. R. (2020). The Cretaceous Period. In The Geologic Time Scale 2020 (Gradstein, F. M., Ogg, J. G., Schmitz, M. D., & Ogg, G. M., Eds.). Elsevier. https://doi.org/10.1016/B978-0-12-824360-2.00027-9
Gore, R. (n.d.), The rise of mammals. In National Geographic. https://www.nationalgeographic.com/science/article/rise-mammals
Ogg, J. G., Hinnov, L. A., Huang, C. (2012a). Cretaceous. In The Geologic Time Scale 2012 (Gradstein, F. M., Ogg, J. G., Schmitz, M. D., & Ogg, G. M., Eds.). Elsevier.
Ogg, J. G., Hinnov, L. A., Huang, C. (2012b). Jurassic. In The Geologic Time Scale 2012 (Gradstein, F. M., Ogg, J. G., Schmitz, M. D., & Ogg, G. M., Eds.). Elsevier.
Jones, P. (2020). Jura Mountains, France/Switzerland. In Around the World in 80 Words. University of Chicago Press. https://doi.org/10.7208/9780226682822-026
National Park Service (2020). Geologic time scale. https://www.nps.gov/subjects/geology/time-scale.htm
Pfiffner, O. A. (2006). Thick-skinned and thin-skinned styles of continental contraction. Special Paper of the Geological Society of America, 414.
Sauquet, H., von Balthazar, M., Magallón, S. et al. (2017). The ancestral flower of angiosperms and its early diversification. Nature Communications, 8. https://doi.org/10.1038/ncomms16047
Scotese, C. R., Song, H., Mills, B. J. W., van der Meer, D. G. (2021). Phanerozoic paleotemperatures: the Earth’s changing climate during the last 540 million years. Earth-Science Reviews, 215. https://doi.org/10.1016/j.earscirev.2021.103503
Wimbledon, W. A. P. (2017). Developments with fixing a Tithonian/Berriasian (J/K) boundary. Volumina Jurassica, XV. https://doi.org/10.5604/01.3001.0010.7467
31 notes
·
View notes
Text
Effigia okeeffeae

By Stolpergeist
Etymology: Ghost
First Described By: Nesbitt and Norell, 2006
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Sauropsida, Eureptilia, Romeriida, Disapsida, Neodiapsida, Sauria, Archosauromorpha, Crocopoda, Archosauriformes, Eucrocopoda, Archosauria, Pseudosuchia, Suchia, Paracrocodylomorpha, Poposauroidea, Shuvosauridae
Referred Species: E. okeeffeae
Status: Extinct
Time and Place: Around 205 million years ago, in the Rhaetian of the Late Triassic
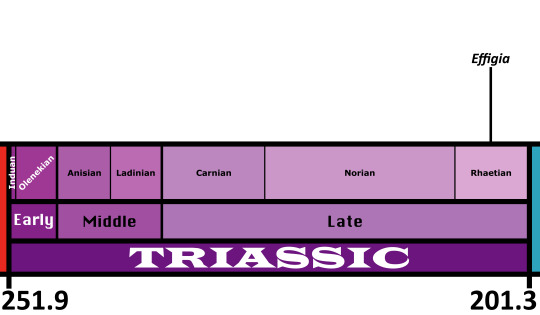
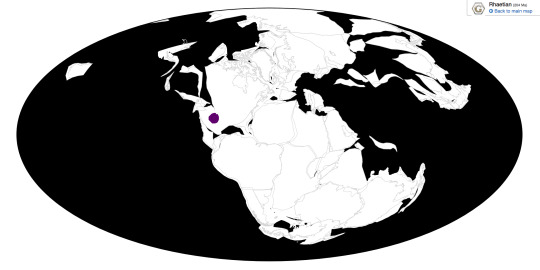
Effigia was found at Ghost Ranch in the Chinle Formation in New Mexico
Physical Description: Effigia was a small, bipedal animal that remarkably resembled dinosaurs it lived with at the time. In total, from head to tail, it would have been about two meters long. It walked on two legs that were held directly underneath the body, and it had small arms that weren’t used in supporting its weight. It had a long body, with a decently sized tail and long neck. The head of Effigia was small and narrow, ending in a beak and having no teeth whatsoever. So, in short, it looked like the later Limusaurus, except it wasn’t feathered - and it wasn’t a dinosaur! This is the first known example of the lightly-built bipedal animal with a beak body plan, aka, the “ostrich” body plan, even though this iteration - the first iteration - had nothing resembling feathers or efficient breathing or hollow bones. In fact, it probably would have breathed primarily based on abdominal muscles, based on its close relatives. And it did have hollow bone walls, much like dinosaurs. Effigia also had a somewhat endothermic body temp - ie, it was closer to warm-bloodedness than modern crocodilians, and may have even been warm-blooded outright. It had five fingers on each hand, though only three of each would have claws; it had four main toes on each foot, unlike the three in theropods, and a little toe raised up (kind of like the fourth toe in theropods). It was rather front heavy, unlike theropods that lean more towards the hip, giving it a forward-leaning appearance.
Diet: Probably herbivorous. While the exact diet of Effigia is murky, the beak of this species indicates it probably would have fed on a variety of plant material, like the later mimics such as Limusaurus and Ornithomimus. However, omnivory was certainly not out of the question.
Behavior: Because Effigia was front-titled, it’s actually not clear whether or not it would have behaved similarly to the animals it resembles. In fact, it doesn’t seem very well adapted for fast movement at all - it looks exactly like the sort of creature that may have tripped over itself on a regular basis. That said, it is entirely possible that it would have balanced itself differently, or adjusted its position in such a way to make up for this oddity in posture. That said, it also had fairly short legs compared to its overall body length, so it’s doubtful that it would have been a fast mover in any position. As such, it probably would have moved slowly throughout its ecosystem, grazing on plants and following fresh vegetation where it came up and utilizing its long neck to reach into areas where food was less accessible, and grabbing on to it for feeding. It may not have been a particularly social animal though, like living archosaurs, it probably would have taken care of its young in some fashion.
Ecosystem: Ghost Ranch was a large floodplain, not quite as forested as the environment had been in earlier times (when they were literally called the “petrified forest”), however, there were still extensive dry forests that experienced dramatic dry and wet seasons each year. These seasons were interspersed with regular flooding, which lead to rapid preservation of a very diverse Late Triassic ecosystem. This was an extremely diverse habitat, with a variety of other reptiles that lived alongside Effigia. There was the slender dinosaur Coelophysis, the weirdly-toothed dinosaur Daemonosaurus, the Silesaurids Kawanasaurus and Eucoelophysis, the Lagerpetid Dromomeron, the early Crocodylomorph Hesperosuchus, the Aetosaur Stenomyti, the phytosaur Redondasaurus, the Drepanosaurs Avicranium and Drepanosaurus, the aquatic archosauriform Vancleavea, the sphenodont Whitakersaurus; coelocanths, ray-finned fish, mystery fish, and even invertebrates such as branchiopods and ostracods. It’s possible that Effigia lived alongside other animals as well, but more research is needed into the exact environment of the Chinle Formation where Effigia was found before that can be confirmed. It is entirely possible that Effigia would have been preyed upon by Coelophysis.
Other: Effigia is so freaking weird, you guys. Like, the Triassic may as well be called the Period in Which Reptiles Tried Out All The Things Dinosaurs Would Later Do (But Only Dinosaurs Would Get Paid For It). It is so similar to later Ornithomimosaurs that at least a few paleontologists used to think that the remains later called Effigia were actually dinosaurs, before their proper reassignment into the Pseudosuchians. This just emphasizes how much different reptiles were trying out new designs and new ideas in the Triassic Period, some of which superficially resembled later dinosaurs - but with surprise twists. It also demonstrates exactly how much crocodile-relatives were diversifying extensively in the Triassic, and how hard they would be hit by the end-Triassic extinction.
~ By Meig Dickson
Sources Under the Cut
Colbert, E. H.. 1947. The little dinosaurs of Ghost Ranch. Natural History 59(9):392-399-427-428.
Heckert, A. B., S. G. Lucas, L. F. Rinehart and A. P. Hunt. 2008. A new genus and species of sphenodontian from the Ghost Ranch Coelophysis Quarry (Upper Triassic: Apachean), Rock Point Formation, New Mexico, USA. Palaeontology 51(4):827-845.
Hunt, A. P., and A. G. Lucas. 1989. Late Triassic vertebrate localities in New Mexico. In S. G. Lucas and A. P. Hunt (eds.), Dawn of the Age of Dinosaurs in the American Southwest, New Mexico Museum of Natural History, Albuquerque 72-101.
Hunt, A. P., and S. G. Lucas. 1993. A new phytosaur (Reptilia: Archosauria) genus from the uppermost Triassic of the western United States and its biochonological significance. In S. G. Lucas and M. Morales (eds.), The Nonmarine Triassic. New Mexico Museum of Natural History and Science Bulletin 3:193-196.
Lucas, S. G., J. A. Spielmann, A. P. Hunt. 2007. Taxonomy of Shuvosaurus, a Late Triassic archosaur from the Chinle Group, American Southwest. New Mexico Museum of Natural History and Science Bulletin 41: 259 - 261.
Lucas, S. G., J. A. Spielmann, and L. F. Rinehart. 2013. Juvenile skull of the phytosaur Redondasaurus from the Upper Triassic of New Mexico, and phytosaur ontogeny. New Mexico Museum of Natural History and Science Bulletin 61:389-400.
Martz, J. W., B. J. Small. 2019. Non-dinosaurian dinosauromorphs from the Chinle Formation (Upper Triassic) of the Eagle Basin, northern Colorado: Dromomeron romeri (Lagerpetidae) and a new taxon, Kwanasaurus williamparkeri (Silesauridae). PeerJ 7: e7551.
Nesbitt, S., M. A. Norell. 2006. Extreme convergence in the body plans of an early suchian (Archosauria) and ornithomimid dinosaurs (Theropoda). Proceedings of the Royal Society B: Biological Sciences 273 (1590): 1045-1048.
Nesbitt, S. 2007. The anatomy of Effigia okeeffeae (Archosauria, Suchia), theropod-like convergence, and the distribution of related taxa. Bulletin of the American Museum of Natural History 302: 84.
Nesbitt, S. J., M. R. Stocker, B. J. Small and A. Downs. 2009. The osteology and relationships of Vancleavea campi (Reptilia: Archosauriformes). Zoological Journal of the Linnean Society 157:814-864.
Nesbitt, S. 2011. The Early Evolution of Archosaurs: Relationships and the Origin of Major Clades. Bulletin of the American Museum of Natural History 352: 1 - 292.
Pritchard, A. C., and S. J. Nesbitt. 2017. A bird-like skull in a Triassic diapsid reptile increases heterogeneity of the morphological and phylogenetic radiation of Diapsida. Royal Society Open Science 4:170499.
Renesto, S., J. A. Spielmann, S. G. Lucas and G. T. Spagnoli. 2010. The taxonomy and paleobiology of the Late Triassic (Carnian-Norian: Adamanian-Apachean) drepanosaurs (Diapsida: Archosauromorpha: Drepanosauromorpha). New Mexico Museum of Natural History and Science, Bulletin 46:1-81.
Rinehart, L. F., S. G. Lucas, A. B. Heckert, J. A. Spielmann, and M. D. Celeskey. 2009. The paleobiology of Coelophysis bauri (Cope) from the Upper Triassic (Apachean) Whitaker quarry, New Mexico, with detailed analysis of a single quarry block. New Mexico Museum of Natural History and Science Bulletin 45:1-260.
Schachner, E. R., P. L. Manning, P. Dodson. 2011. Pelvic and hindlimb myology of the basal archosaur Poposaurus gracilis (archosauria: Poposauroidea). Journal of MOrphology 272 (12): 1464 - 1491.
Schaeffer, B. 1967. Late Triassic fishes from the western United States. Bulletin of the American Museum of Natural History 135(6):285-342.
Sues, H.-D., S. J. Nesbitt, D. S. Berman and A. C. Henrici. 2011. A late-surviving basal theropod dinosaur from the latest Triassic of North America. Proceedings of the Royal Society B 278:3459-3464.
#Effigia#Effigia okeeffeae#Poposaur#Pseudosuchian#Triassic#Archosaur#Reptile#Triassic Madness#Palaeoblr#Triassic March Madness#Prehistoric Life#Paleontology#Prehistory
198 notes
·
View notes
Text
Thecodontosaurus antiquus

By Scott Reid
Etymology: Reptile with Teeth in Sockets
First Described By: Lydekker, 1890
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Sauropodomorpha, Bagualosauria
Status: Extinct
Time and Place: Between 208 and 201 million years ago, in the Rhaetian age of the Late Triassic
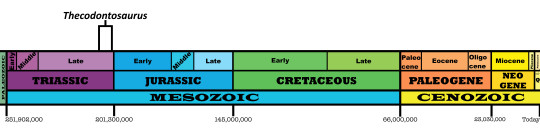

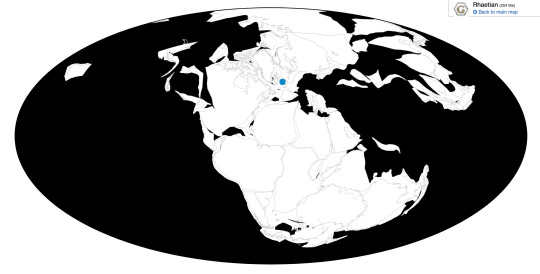
Thecodontosaurus is known from the Magnesian Conglomerate Formation, the Microlestes Quarry, and the Tytherington Quarry of Southwestern England
Physical Description: Thecodontosaurus was a small “prosauropod” - the informal group of the dinosaurs more closely related to sauropods than to other dinosaurs, but aren’t sauropods themselves. These dinosaurs, while rather plain on the outside, are notable in a few ways - the biggest ones being that they are the most common dinosaurs of the Triassic, and that they show the evolutionary path from “generic dinosaur” to “giant behemoth sauropod”. Thecodontosaurus falls on the earlier end of that scale, being a small, bipedal, lightweight animal. It was only about two meters long, and less than a meter high at the hips. It had a fairly short neck with a larger head on the end than is seen in later sauropodomorphs (where necks got longer, but heads got smaller); it had many small, leaf-shaped teeth in its mouth, and large eyes in its skull. It had long and narrow hands, with very short front limbs and longer hind limbs; its tail was especially long, longer than the rest of the body put together. Some individuals were smaller than others, probably because they lived on a separate island and experienced insular dwarfism. Given its extremely small size - only weighing 11 kilograms - it was covered in fluffy protofeathers to aid in thermoregulation.

By Slate Weasel
Diet: With the serrated leaf-shaped teeth, Thecodontosaurus was definitely an herbivore, and a low browser at that. This is significant, because Prosauropods started out as Carnivores, and then transitioned to herbivory; Thecodontosaurus was, thus, well into that transition.
Behavior: Thecodontosaurus was probably a fairly social animal, having been found in decent numbers in concentrated locations. As such, small flocks of these dinosaurs would probably have roamed the coastline and scrub-caves. They would have probably had to be at least somewhat skittish, really only having their own speed to get away from predators. They also would have been quite active and warm-blooded. Thecodontosaurus would probably have spent most of its time eating, but it also probably took care of its own young and brought food back to the babies.

By José Carlos Cortés
Ecosystem: Thecodontosaurus lived during an interesting time in Earth’s History - the Rhaetian Age, though right before the Triassic extinction and probably the best time to look at how the unique life of the Jurassic managed to slip through said event, is not well-represented in the fossil record. Thus, Thecodontosaurus and its environs are, while sparse, uniquely important in terms of understanding the transition of Mesozoic life. This was a series of small islands called the Mendip Archipelago, filled with mosses, cycads, and algae, with frequent caves lining the environment. As such, many kinds of fish were especially common here, including the weird Palaeonisciforms, sharks, and more ray-finned fish. Some early almost-mammals were also present, such as Kuehneotherium and Thomasia, as well as Eozostorodon. There were many other reptiles as well, though very few were dinosaurs; many were just lizardy-things. There was the mysterious Palaeosaurus, the phytosaur Rileya, and the Rhynchocephalian Diphydontosaurus. As for other dinosaurs, there may have been theropods to prey on Thecodontosaurus, but its only major dinosaur neighbor seems to have been another prosauropod, Asylosaurus.

By Ripley Cook
Other: Thecodontosaurus, in addition to being an important step in the sequence from basic dinosaurs to the sauropods, is a notable discovery for being one of the first dinosaurs found. It was actually the fifth (non-avian) dinosaur named, after Megalosaurus, Iguanodon, Streptospondylus, and Hylaeosaurus. It has, thus, been used as the name of many dinosaurs around the world, but very few of these finds are actually Thecodontosaurus.
~ By Meig Dickson
Sources under the Cut
Apaldetti, C., R. N. Martinez, O. A. Alcober and D. Pol. 2011. A new basal sauropodomorph (Dinosauria: Saurischia) from Quebrada del Barro Formation (Marayes-El Carrizal Basin), Northwestern Argentina. PLoS ONE 6(11):e26964:1-19
Benton, M. J., L. Juul, G.W. Storrs and P.M. Galton, 2000, "Anatomy and systematics of the prosauropod dinosaur Thecodontosaurus antiquus from the upper Triassic of southwest England", Journal of Vertebrate Paleontology 20(1): 77-108
Galton, P. M. 2007. Notes on the remains of archosaurian reptiles, mostly basal sauropodomorph dinosaurs, from the 1834 fissure fill (Rhaetian, Upper Triassic) at Clifton in Bristol, southwest England. Revue de Paléobiologie 26(2):505-591.
Galton, P.M., Yates, A.M., & Kermack, D. (2007). "Pantydraco n. gen. for Thecodontosaurus caducus Yates, 2003, a basal sauropodomorph dinosaur from the Upper Triassic or Lower Jurassic of South Wales, UK". Neues Jahrbuch für Geologie und Paläontologie, Abh., 243: 119-125
Griffin, C. T., and S. J. Nesbitt. 2016. The femoral ontogeny and long bone histology of the Middle Triassic (?late Anisian) dinosauriform Asilisaurus kongwe and implications for the growth of early dinosaurs. Journal of Vertebrate Paleontology 36(3):e1111224:1-22
Haughton, S.H., 1924, "The fauna and stratigraphy of the Stormberg Series", Annals of the South African Museum 12: 323–497
Langer, M. C., M. D. Ezcurra, J. S. Bittencourt and F. E. Novas. 2010. The origin and early evolution of dinosaurs. Biological Reviews 85:55-110
McPhee, B. W., J. N. Choiniere, A. M. Yates and P. A. Viglietti. 2015. A second species of Eucnemesaurus Van Hoepen, 1920 (Dinosauria, Sauropodomorpha): new information on the diversity and evolution of the sauropodomorph fauna of South Africa's lower Elliot Formation (latest Triassic). Journal of Vertebrate Paleontology 35(5):e980504:1-24
Morris, S. December 13, 2013. Thecodontosaurus, Bristol's own dinosaur, to go on display at university. The Guardian.
Naish, D., and D. M. Martill. 2007. Dinosaurs of Great Britain and the role of the Geological Society of London in their discovery: basal Dinosauria and Saurischia. Journal of the Geological Society, London 164:493-510
Nesbitt, S. J.; Barrett, P. M.; Werning, S.; Sidor, C. A.; Charig, A. J. (2013). "The oldest dinosaur? A Middle Triassic dinosauriform from Tanzania". Biol. Lett. doi:10.1098/rsbl.2012.0949.
Novas, F. E., M. D. Ezcurra, S. Chatterjee and T. S. Kutty. 2011. New dinosaur species from the Upper Triassic Upper Maleri and Lower Dharmaram formations of Central India. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 101:333-349
Otero, A., and D. Pol. 2013. Postcranial anatomy and phylogenetic relationships of Mussaurus patagonicus (Dinosauria, Sauropodomorpha). Journal of Vertebrate Paleontology 33(5):1138-1168
Riley, H., and S. Stutchbury. 1836. A description of various fossil remains of three distinct saurian animals discovered in the autumn of 1834, in the Magnesian Conglomerate on Durdham Down, near Bristol. Proceedings of the Geological Society of London 2:397-399
Smith, N. D., and D. Pol. 2007. Anatomy of a basal sauropodomorph dinosaur from the Early Jurassic Hanson Formation of Antarctica. Acta Palaeontologica Polonica 52(4):657-674
Upchurch, P., P. M. Barrett, and P. M. Galton. 2007. A phylogenetic analysis of basal sauropodomorph relationships: implications for the origin of sauropod dinosaurs. In P. M. Barrett, D. J. Batten (eds.), Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs, Special Papers in Palaeontology 77:57-90
Vickers-Rich, P., Rich, T.H., McNamara, G.C. & Milner, A. (1999). "Agrosaurus: Australia's oldest dinosaur?". Records of the Western Australian Museum Supplement 57: 191-200
Weishampel, David B; et al. (2004). "Dinosaur distribution (Late Triassic, Europe)." In: Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.): The Dinosauria, 2nd, Berkeley: University of California Press. Pp. 521–525
Whiteside, D.I. and Marshall, J.E.A. (2008) "The age, fauna and palaeoenvironment of the Late Triassic fissure deposits of Tytherington, South Gloucestershire, UK". Geological Magazine, 14(1): 105-147
Xing, L., T. Miyashita, J. Zhang, D. Li, Y. Te, T. Sekiya, F. Wang and P. J. Currie. 2015. A new sauropod dinosaur from the Late Jurassic of China and the diversity, distribution, and relationships of mamenchisaurids. Journal of Vertebrate Paleontology 35(1):e889701:1-17
Yates, A. M. (2003). "A new species of the primitive dinosaur Thecodontosaurus (Saurischia: Sauropodomorpha) and its implications for the systematics of early dinosaurs". Journal of Systematic Palaeontology 1(1): 1-42
Yates, A.M. & Kitching, J. W. (2003). "The earliest known sauropod dinosaur and the first steps towards sauropod locomotion". Proc. R. Soc. Lond.: B Biol Sci. 2003 Aug 22; 270(1525): 1753–8
#Thecodontosaurus antiquus#Thecodontosaurus#Prosauropod#Sauropodomorph#Dinosaur#Prehistoric Life#Paleontology#Prehistory#Palaeoblr#Factfile#Mesozoic Monday#Dinosaurs#Eurasia#Triassic#Herbivore#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature
229 notes
·
View notes
Text
More Things in Heaven and Earth
There are more things in heaven and earth, Horatio,
Than are dreamt of in your philosophy.
- Hamlet
Perhaps, in this age of instantaneity, I am a bit late to the party, but the recent announcement regarding the Trappist-1 system has been in my thoughts. These have thence wandered towards more general topics in life and evolution, and I feel the urge to write.
Although I encourage anyone with an interest to check primary sources on this system (www.trappist.one, operated by the original discovery team, is very useful) I will present some facts salient to my discussion here.
The Trappist-1 system is composed of at least seven planets orbiting a small, ultra-cool star (about the size of Jupiter) less than 40 light years from earth. Ultra-cool stars like Trappist-1a are the most common stars in the galaxy, and compared to other types of stars, allow for the easier detection of their planets and better determination of their planets’ qualities. In the case of Trappist-1, Michael Gillon et al. determined that all seven planets were rocky, earth-sized, with three being capable of supporting liquid water – and thus life as we know it.
It seems difficult to know whether these planets might support life: Given the right conditions and enough time, the development of life in some form is likely inevitable (this statement is really just a truism) – the issue is knowing what the right conditions are. Whereas the last few hundred years of work in biology have yielded a reasonable understanding of life on Earth, I submit that our understanding of Life, in general, is poor. Surely, given the vastness of space, life must exist elsewhere in the universe – ours cannot be the only light flickering in the dark. Life on Earth, then, is only a single specific case of a broader phenomenon, and drawing general conclusions from a single specific case is always dubious!
I have no doubt that the discovery of extra-terrestrial life, if and when it ever occurs, would be the most significant, paradigm shattering event in our understanding of life since Darwin published Origin of Species. I also have no doubt that we cannot know what to expect. Life is shaped by its environment and Earth’s, throughout its history of life, has remained within a relatively narrow band of stability, close enough to what we see today. And yet, which of us, without foreknowledge, might imagine an insect, an anenome, or a sea sponge? How wondrous are the forms that have existed only on this planet! Think of the prokaryotes in their underappreciated, dominant, splendor. Look at the fauna of the Cambrian “explosion” – Anomalacaris, Opabinia, Marella, Hallucigenia – and see how long it took us to resolve their anatomy. Behold the Tully Monster and the time and effort it took to discern even that it might be a vertebrate – a conclusion which remains controversial! The last one hundred years of palaeontology have yielded us more aliens than a millennia of astronomy. In time, life adapts to its conditions – yet all this diversity branches from a single point of life on a single planet. Ours is a tidal pool of stability on the edge of a vast ocean of possibility. Surely there are more things in heaven then can be dreamt about in our philosophy.
I can think of only one universal truth which must define the possibilities of life – that of natural selection. The notion of “survival of the fittest” isn’t that revolutionary – some (even Ann Coulter!) have noticed that it’s a syllogism. It’s true simply by its definitions. The fit survive, because the fit are defined as those that survive. Thus, whereas this statement is a poor epitome of Darwin’s work, it serves well as an introduction to the universal law of natural selection. As Dawkins explains in The Selfish Gene (which, despite my initial biases, I recommend most highly – Dawkins exposits the following better and in more detail) “survival of the fittest” is a special case of the law of “survival of the stable” – which is true by definition. Dawkins writes, “the Universe is populated by stable things.” Atoms, molecules, mountains, and planets all exist because they are stable. Self-replication of molecules – the basic, key mechanism of Life in general -- throws a wrench in the works. Any stable self-replicating molecule, in a rich soup of its building blocks, quickly becomes ubiquitous. As the building blocks become rare, any self-replicating molecule with a slight difference allowing it to make more efficient use of its building blocks, or steal from other self-replicating molecules, or prevent theft from itself (I anthropomorphize for simplicity and don’t mean to imply agency); or giving it any slight advantage would soon become more common than its peers. From this basis of replicating molecules (which, on Earth, are DNA) and accumulation of slight advantages, it is not such a large leap to the development of many and varied forms of Life. All Life requires is a self-replicating molecule, and the algorithm of natural selection takes care of the rest. The revolution of Darwin’s work was recognizing this algorithm in life on Earth, evincing it, and resolving its implications. The revolution of The Selfish Gene as the recognition that the gene, as composed of self-replicating molecules, is the unit of evolution, rather than the individual or species.
So, whereas we cannot predict at all what life in the universe might look like, we can be certain that it has been shaped by the universal algorithms of stability and natural selection (Darwin’s enduring legacy) and that all that life requires is a replicator (Dawkins’ enduring legacy).
I really don’t know how to wrap this up, so I will finish with an aside. In The Selfish Gene, Dawkins demonstrates convincingly that genes, because of the universal algorithm of natural selection, act selfishly through the individual to promote their own propagation. Selfish genes are better at surviving, so only the selfish genes survive. In nature, therefore, everything is selfish – altruism exists only when it promotes the propagation of the genes. Humanity is unique because of consciousness – Dawkins writes that “We, alone on Earth, can rebel against the tyranny of the selfish replicators.”
And so we have. I have an example – one that formed the basis for this essay: when we discovered Trappist-1. A team worked together, with themselves and with colleagues internationally, not for selfish interests, but for a noble goal bigger than any one person.
Humanity has defied, and we preach defiance to the stars.
1 note
·
View note
Text
The evolution of biodiversity: ever-increasing or did it hit a ceiling?
By Gareth Willmer
Preserving biodiversity is one of the key debates of our time – but another subject of hot debate in recent decades among evolutionary experts is how biodiversity has changed over the past few hundred million years. New findings are challenging the conventional view on this.
Increasing knowledge on historical patterns of biodiversity could also help us understand animals’ ability to adapt and what climate change means for evolution.
The traditional view is that species have increased in diversity continuously over the past 200 million years, particularly in the last 100 million, leading to more diversity now than ever before. But some recent studies suggest biodiversity has tended to stay largely the same, with only occasional surges.
‘Our findings strongly contradict past studies that suggested unbounded diversity increases at local and regional scales over the last 100 million years,’ said a fresh study on terrestrial species. It found no evidence of a rise in diversity in the past 66 million years, following a brief two- to three-fold increase over a couple of million years after the mass extinction of the dinosaurs at the end of the Cretaceous Period and as mammals began to thrive.
The story was similar for a study on marine species, with the researchers finding little change in ocean biodiversity over the past 200 million years.
‘We find that ecosystems are relatively stable in terms of how much diversity they have over tens to hundreds of millions of years,’ said Professor Richard Butler, a palaeobiologist at the University of Birmingham, UK, who worked on both studies as part of the TERRA project. ‘Rather than seeing exponential increases, it’s more like a short-term quite dramatic increase and then a relatively flat line.’
The findings imply that diversity is capped at a certain limit and that species numbers are ultimately limited by factors such as the availability of resources like water and space, says Prof. Butler. Various researchers have been moving towards this view for some time, he says, but new approaches like those adopted in the TERRA project have helped bolster the evidence.
Inflation
Prof. Butler explains that the changing picture is being revealed by methods that help reduce the long-standing challenge experts have faced of biases in the fossil record. The issue is that the higher abundance of preserved fossils in the more recent past leads to artificial inflation of diversity.
One of their approaches has been to take a regional view rather than a global one. ‘There’s been a tradition in the field that what we’re interested in is trying to understand global biodiversity… but as you go deeper in time, you might have whole continents that aren’t sampled (as no fossils have been found) for some time intervals,’ said Prof. Butler.
The researchers built on data collected over the past 20 years in the Paleobiology Database, which gathers global taxonomic data for organisms throughout geological history. They then created algorithms to sample similar-sized areas where fossils had been found through time.
Using these methods, their studies highlighted significant variations in diversity at different locations across time – such as it tending to be much higher at the equator than at the poles – emphasising the need for a regional view to get the full picture.
The researchers have also started working to integrate their results with the data of climate modellers. They want to see how climate shifts have affected the distribution of biodiversity over deep time, which they are doing by looking at the types of conditions that prevailed globally and in different parts of the world when certain animals were around.
Prof. Butler notes, however, that estimating the level of biodiversity from the fossil record is still challenging given the many gaps in the database. For example, teeth are much harder to use for distinguishing dinosaur than mammal species.
‘Whether life diversified (continuously) by way of numerous jumps and steps… or in fact reached a kind of full capacity and then didn’t shift much is still hotly debated.’
Professor Michael Benton, University of Bristol, UK
Trends
The complexity of teasing out historical biodiversity trends is emphasised by the different picture found in the past studies of Dr Hélène Morlon, an evolutionary ecologist at the French National Centre for Scientific Research.
Her results suggested no set limit on biodiversity, unlike the work of TERRA, but slowdowns in the rate at which new species emerge as ecological niches or habitats become more crowded over geological time. ‘We found models where diversity is not bounded – with a fixed number of species that doesn’t change – and it’s not exponentially increasing, but there’s this type of slowdown in the way that diversity accumulates,’ she said.
Dr Morlon has found further evidence for such slowdowns as part of a project she leads called PANDA, which has sought to develop better models and tools for harnessing data in evolutionary, or phylogenetic, trees to study diversification at a finer scale. Among its results, the researchers have identified five main diversification patterns among vertebrates, providing a framework for understanding the evolution of biodiversity in deep history.
They also had a surprise result. They found evidence that cooler periods over the past 66 million years coincided with faster rates of body size evolution in birds and mammals, even though they found that overall diversification tends to slow in cooler climates.
The researchers suggest that this result could be because body sizes have a specific relationship to temperature or, more widely, that certain traits might be decoupled from diversification. All this suggests that further research is needed on the links between the evolution of body traits and diversification.
‘The link between different levels of evolution is super complicated and not well-known,’ said Dr Morlon. ‘We need pretty complicated models that are not that easy to develop.’
But while there is still much to unravel, developments in this area may apply to modern-day issues too.
While researchers emphasise that past evolutionary trends are not directly translatable to today because of the vastly different timescales, they say studies could provide some clues about the ability of species to adapt and the combinations of characteristics such as feeding patterns or size that put species at risk. This research could also provide implications for how human-driven climate change might affect future evolution.
‘We can estimate at these timescales what happens when you change temperature by that many degrees – what’s the effect on evolutionary rates?’ said Dr Morlon. ‘It also at least gives perspective to understand how biodiversity might answer to current changes.’
Diversification
Meanwhile, the debate about diversification continues.
Professor Michael Benton, who studies vertebrate palaeontology at the University of Bristol, UK, has been a strong proponent of continually rising biodiversity and still leans towards that view.
While agreeing that the fossil record is incomplete, he says the broad outline of the order of fossils is not misleading and broadly documents the ‘rise and fall of major groups through time.’
Prof. Benton cites the example of the proliferation of birds to today’s 10,000 species. To suggest that biodiversity has remained constant since birds came into being is like suggesting that bird species replaced a similar number of pre-existing species of flying animals, he says.
Similarly, he said: ‘There was a time when animals didn’t form reefs or burrow under the seabed; then those innovations emerged and this presumably allowed a net increase in biodiversity’.
He agrees, however, that the answers remain far from clear cut and much more work is needed to get to the bottom of it – but that the many advances in computational tools give a big helping hand.
‘It still seems to be unresolved to what extent the fossil record gives a reasonable picture of life,’ said Prof. Benton. ‘Whether life diversified (continuously) by way of numerous jumps and steps… or in fact reached a kind of full capacity and then didn’t shift much is still hotly debated.’
The research in this article was funded by the EU. If you liked this article, please consider sharing it on social media.
Published by Horizon
source https://horizon.scienceblog.com/1290/the-evolution-of-biodiversity-ever-increasing-or-did-it-hit-a-ceiling/
0 notes
Text
The Science of Jurassic Park & More: Part Three
In today’s article, we will be continuing the final part of our deep-dive into the world of palaeontology with Doctor David Button from London’s Natural History Museum.
In Part Two, David talked about his favourite dinosaurs, whether we are still discovering new dinosaurs, and much more. If you missed it, click here to take a read.
Many of the images in the article ahead are also courtesy of our friends at Jurassic Vault – so go show them some love if you haven’t already!
Without any further ado, let’s kick off our final part to this series by talking about the accuracy of the fan favourite Spinosaurus!
Image from Jurassic Park Wiki.
How accurate was the Spinosaurus we saw in Jurassic Park 3 (2001) at the time of release?
The Spinosaurus of Jurassic Park III was actually reasonably accurate at the time of release, as it was not well known at the time. Its main problem was the orientation of the hands – as with most of the other theropods in the franchise, it was depicted with permanently pronated “bunny hands”, even though theropods were unable to pronate their hands into such a position.
How do you see/research intelligence of extinct animals that died millions of years ago?
We can get an idea of the cognition of an extinct animal by taking a cast of the interior surfaces of the braincase – producing what is known as an endocast. This used to be performed by injecting latex into a dinosaur braincase and then cutting it open. Fortunately, however, these days we are able to produce these digitally by CT-scanning dinosaur braincases. The endocast gives us a replica of the shape of the brain and surround soft tissues.
From this, we can examine the shape of the brain and its constituent lobes to get an idea of the sensory abilities of an animal, and from the dimensions of the inner ear we can even calculate the range of sounds an animal could hear. The overall size and shape of the brain also give us a rough idea of the animal’s intelligence relative to that of other creatures. We can then also compare this with other evidence of dinosaur behaviours, such as associations of skeletons and footprints, to get an idea of their cognition and behavioural complexity. However, it should be stated that brain size does not necessarily correlate tightly with intelligence, and certainly many animals with small brains are still capable of complex cognition and behaviours. Consequently, we can only get a vague impression of just how smart dinosaurs may have been.
Would a Mosasaurus have slept in the same way a Whale sleeps?
Sleeping poses a problem for marine animals that need to come up for air, but cannot haul out onto land. Whales have solved this by only putting part of the brain to sleep at a time, still allowing them to surface. Sea turtles, however, have different solutions. Sometimes they sleep floating at the surface. Otherwise, however, sea turtles are able to exploit their low metabolisms. They slow their metabolic rate and oxygen use so as to not require another breath for hours, allowing them to sleep for that time under the surface.
It is difficult to say for sure Mosasaurs would have slept. However, there is evidence that Mosasaurs were warm-blooded, with a relatively high metabolic rate. Consequently, I think they would have been more likely to sleep like a whale than like a turtle.
Stiggy.....is it a growth phase of the pachycephalosaur?
Stygimoloch, Dacrorex and Pachycephalosaurus are differently-sized genera of pachycephalosaur, all known from the same time and place. Work on the bone histology of the animals has shown that the known specimen of Dacrorex is an infant, Stygimoloch a juvenile, and Pachycephalosaurus an adult. This suggests that they may all be growth phases in the life of a single animal. However, it may equally mean that they are three different animals that just happen to be known from different life stages. Unfortunately, pachycephalosaur remains are generally rare, and with such small sample sizes it is difficult to satisfactorily prove this problem one way or the other.
However, that being said, it seems more likely to me that these dinosaurs are all different life stages of the same animal than that there were three very similar pachycephalosaurs living in the same time and place. Hence, I think it is best to assume this option, until more specimens let us test this more thoroughly. I do admit that this is disappointing, though, as Stygimoloch was one of my favourite dinosaurs.
Is it true that we don’t know what Carnotaurus’ feet look like? And it’s tail?
It is true that specimens of Carnotaurus that we do have do not preserve these parts of the skeleton, so we do not know exactly what they looked like. However, Carnotaurus was an Abelisaurid, and these parts of the skeleton are known from other Abelisaurs, such as Majungasaurus, allowing us to fill them in with a reasonable degree of accuracy. This suggests that the tail of Carnotaurus would have been rather unremarkable. The hind legs would have been quite stocky, but what we do have from Carnotaurus indicates that its legs would have been relatively longer than those of Majungasaurus, and so Carnotaurus would have been a more competent pursuit predator.
The question I always wanted to ask is. How come they decided to go for reptiles instead of birds for their Jurassic Park films and why are the raptors are bigger than their real-life counterpart?
Jurassic Park originally used reptiles as a model for dinosaurs partially as this was the prevailing view when the book was first written, and also as that was the public expectation of dinosaurs. Nonetheless, the book and first movie movie did draw heavily on birds for some of the depicted behaviours, so that each were quite radical when they came out. Since then, however, the Jurassic Park franchise has stuck with more reptilian dinosaurs despite the science leaving that far behind. This is probably partially to preserve continuity, and partially because the public still, unfortunately, expects and prefers their dinosaurs to be as reptilian as possible when watching movies.
Velociraptor was originally scaled-up to more resemble the size of Deinonychus by Michael Crichton when he was writing Jurassic Park. This partially followed the unusual classification of Dromaeosaurids by the palaeontologist Greg Paul, who considered Velociraptor mongoliensis and Deinonychus antirrhopus to both belong to the same genus. Jurassic Park followed this, featuring Deinonychus, but calling it Velociraptor Antirrhopus. This also explains why the featured Velociraptor skeleton was discovered by Grant in the USA, as opposed to Mongolia, where Velociraptor actually lived. However, no other palaeontologists agree with this classification, not the least because the two animals lived on separate continents, 30 million years apart! Nonetheless, Crichton decided to stick with the name Velociraptor, because he considered it more dramatic.
When it came to adapting Jurassic Park for film, the raptors were increased in size again. This was due partially to Spielberg, reportedly, being unimpressed with the size of Deinonychus. However, it was also due to the discovery of Utahraptor at around the same time – it had not yet been described and named, but there were reports of large raptor bones being found at the time that inspired the production crew of the movie.
If an eccentric millionaire was able to bring back dinosaurs and make a Jurassic Park, what is the probability of them breaking out and creating all the chaos seen in the films?
That’s a big if! However, if they did, I am sure that dinosaurs would break out every so often. How much chaos were then able to cause would be directly down to how well-prepared the millionaire had been. If they had stringent systems in place, an escaped dinosaur should be quickly containable – although their large size and strength would pose significant challenges, they would not be any more vicious or intelligent than other zoo animals. However, if they had under prepared with inadequate staff, and over reliance on untested automated systems and untrustworthy employees, then, well…
Considering how obvious this is, and I've asked this before, does the don't move thing for Tyrannosaurus sound ridiculous and is 12.5 meters the plausible maximum for the king of the Cretaceous? If a Rex went 32 mph, would it trip over?
You are correct in that vision-based eyesight for a large predator like Tyrannosaurus makes no sense at all. I am glad the series has been downplaying this since Jurassic Park.
How quickly Tyrannosaurus could run remains a strong point of contention within palaeontology. Still, most estimates do hover around the region of a running speed of 11-13 metres per second (~25 mph) – this seems sensible to me. At higher speeds the problem is not that Tyrannosaurus would necessarily fall over, but that if it did happen to trip, the head would hit the ground with such momentum that it would be fatal. Hence, it is possible that it could have briefly used such a burst of speed when closing in on prey, but it would not have used such speeds during normal locomotion or pursuit.
Are Giganotosaurus and Carcharodontosaurus considered Allosaurid or Allosauroid? And if suddenly someone brings dinosaurs to life through their original DNA, would you use frog or bird DNA, to make them as accurate as possible? And is the Jurassic Park T-Rex a bit shoehorned and shrinkwrapped?
Carcharodontosaurids such as Giganotosaurus and Carcharodontosaurus itself are Allosauroids. However, they are not also Allosaurids, but are instead more closely related to Neovenatorids: these two groups together form the Carcharodontosauria.
Were it possible to extract DNA from dinosaur fossils to revive them, I would certainly use bird DNA to plug the gaps. This would both make the resulting dinosaurs more accurate and also, due to using a more closely related animal, make it more likely that the created embryos would be viable in the first place. Unfortunately, though, it does not appear possible for DNA to survive for that long in fossils.
‘Shrink-wrapping’ is a term used to describe palaeoart that does not put enough flesh and other soft tissues on a dinosaur’s bones. As you say, the Tyrannosaurus in Jurassic Park has suffered a little bit of shrinkwrapping – the spine, shoulder blade, ribs and hips are very obvious, as are some of the edges of the skull. However, overall, most of the dinosaurs in Jurassic Park are not too bad for shrinkwrapping – the Tyrannosaurus, for example, just looks underfed – this may be as the need to create a moving 3D model means the animators will get some kind of feel for the volumes of muscle that should be present. Still, most of the dinosaurs should have lips, and the Pterosaurs in Jurassic World were shrinkwrapped to a horrifying degree, as were the heads of the Apatosaurs in the same movie.
And which parts of the Jurassic Park T-Rex would you rather fix to give nod to accuracy?
I would draw upon evidence of the integument of tyrannosaurs to make it both more accurate, but also more visually interesting. Although we know that Tyrannosaurus was mostly covered in scales, we also know that it had some regions of naked skin (and, possibly, some feathers). Furthermore, we know that Tyrannosaurus’ face was armoured by large, overlapping scales – it would be nice to include these. Despite this, I do also think that Theropods such as Tyrannosaurus would have had lips, and so would include those. The ridges on Tyrannosaurus’ head were probably for display, and it would have had good colour vision, so brightly colouring those seems both plausible and visually interesting.
In addition, drawing on the above discussion about ‘shrinkwrapping’, I would beef Tyrannosaurus up a bit, especially around the legs and tail (both related to the leg muscles, but also likely fat stores in the tail as seen in modern reptiles). I tend to like my dinosaurs to be bulky, as we tend to estimate volumes of soft tissue in animals in general from their skeletons.
How much can you rely on fused bone sutures to understand a dinosaur’s growth stage? For example, why does a juvenile T. rex have fused sutures but appears to be young when looking at its histology?
The degree of sutural fusion in a dinosaur is often used to get a rough idea of life stage although, as you say, there are many caveats. In tyrannosaurs, for example, sutures in the axial and appendicular regions of the skeleton fused very early during growth, whereas many of those in the skull never fused, even in adulthood. The fusion of sutures between bones is governed, in part, on the forces acting upon them. Consequently, the stresses place on the tyrannosaur’s back and limbs could cause them to fuse early. By contrast, the open sutures between tyrannosaur facial bones served to help absorb and redistribute shocks encountered during feeding.
With those caveats in mind, bone histology remains the best tool to understand whether a dinosaur had reached adulthood. Still, unfused sutures in certain bones are still useful in showing us whether a dinosaur was still a juvenile, even if their fusion may not necessarily indicate that they were yet an adult. For example, the unfused scapulacoracoid of the mounted skeleton of Giraffatitan means that we know it was still growing, and was not fully adult.
Even though modern evidence suggests that adult tyrannosaurs probably had no feathers, does evidence suggest that young ones may have had feathers — possibly to help regulate heat?
We know that ancestral Tyrannosauroids – such as Dilong and Yutyrannus had feathers, and so they were secondarily lost in Tyrannosaurids, perhaps due to their large size obviating the need for insulation (which, indeed, would have caused heat stress in such large animals). Consequently, I think it is very plausible that tyrannosaur chicks would have had some kind of down-like coat to help keep them warm, especially in more northerly regions of tyrannosaur habitat. Indeed, it seems probable that the small, polar Tyrannosaurid Nanuqsaurus would have retained a feathery coat in adulthood, to help it last the cold winters.
It should also be noted that, although preserved skin impressions show that Tyrannosaurids such as Tyrannosaurus and Albertosaurus were predominately scaly, we still cannot rule out the presence of small regions of feathers, whether they were relicts of their evolutionary history or growth, or even for display.
What do we know of the lifespan of dinosaurs? Elephants have a gestational period of two years. Do we know anything about length of egg carrying or incubating time? Massive dinosaurs (Titanosaurs: how long until they reach adulthood? Would it be safe to assume they lived a long time?
We can get an idea of the age of a dinosaur specimen from looking at sections of its bones, especially the long bones of the limbs. In many cases, the bones of dinosaurs bones show growth rings – like those of trees – which can be counted to establish an age. Unfortunately, some dinosaurs – especially whilst young – grew too quickly to leave rings. In other cases, remodeling of bone structure after reaching adulthood also overpinted the rings. However, even then, the texture of bone indicates how rapidly the animal was growing when it died. Comparing these between differently-sized individuals of a dinosaur species allows us to establish growth curves. This, in turn, helps to establish when a dinosaur reached sexual maturity – as indicated by a slowdown in growth as resources are diverted for reproduction – and also the age at which maximum body size was achieved.
These results show that most dinosaurs grew fast – even large dinosaurs reached sexual maturity in their teens, and maximum body size in their twenties. Maximum lifespan is harder to establish, though, as it is impossible to verify whether any particular dinosaur died of old age. However, we know of some individuals that were at least 38 years old. However, it should be noted that very few dinosaurs would have reached old age, or even full body size – most would have died relatively young, due to their violent lifestyles. Once reaching full body size, however, they would be relatively safe from attack, and maximum lifespans of 50 years or more for large dinosaurs would seem likely.
Incubation times of dinosaur eggs can be calculated by counting growth rings in the teeth of dinosaur embryos. These indicate that incubation times in dinosaurs were variable. Most dinosaur eggs probably incubated for similar timescales to those seen in modern reptiles, taking from between 3-6 months to hatch. However, those of more birdlike dinosaurs, such as Troodon, hatched more quickly (but still more slowly than in modern birds), after only ~70 days.
Even though this means Titanosaur eggs may have taken several months of burial to hatch, the low investment a mother would have to put into each egg means that she would be able to lay multiple clutches per year, each containing as many as 30 eggs, for an annual total of perhaps as many as 200 eggs in the largest species. This is a much higher reproductive rate than a modern elephant, which has to invest 22 months of pregnancy, followed by 3-5 years of devoted parental care in a single calf. The lesser pressures of egg-laying relative to pregnancy are one reason why dinosaurs were able to reach much larger sizes than mammals such as elephants.
If the Baryonyx was around today would we see them in the swamps of Florida and or Louisiana?
Evidence from the isotopic composition of the bones of Spinosaurids such as Baryonyx, as well as from fish scales preserved in the animal’s stomach, indicate that it mostly ate fish and spent a large amount of its time in or around water. The warm, swampy environment of the Everglades would seem like ideal habitat for Baryonyx – and, indeed, is not too far from the wet environment of the Wealden Formation from which it is known. No Spinosaurids are currently known from North America, but that does not mean that Baryonyx would not be able to thrive were it somehow introduced there!
We really hope you found this three-piece series interesting! Make sure you also check out Part One and Part Two.
A massive thank you, again, to David for working on this series with us. We’ve found it incredibly interesting – and we hope you have too! Make sure to follow David on Twitter if you aren’t already, and stay tuned to The Jurassic Park Podcast for all the latest Jurassic Park news!
Written by:
Tom Fishenden
#article#doctor david button#david button#ask a palaeontologist#ask a paleontologist#paleontologist#paleontology#science of jurassic park#tom fishenden#tom jurassic
0 notes
Quote
On the one hand, Orientalism acquired the Orient as literally and as widely as possible; on the other, it domesticated this knowledge to the West, filtering it through regulatory codes, classifications, specimen cases, periodical reviews, dictionaries, grammars, commentaries, editions, translations, all of which together formed a simulacrum of the Orient and reproduced it materially in the West, for the West. The Orient, in short, would be converted from the personal, sometimes garbled testimony of intrepid voyagers and residents into impersonal definition by a whole array of scientific workers. It would be converted from the consecutive experience of individual research into a sort of imaginary museum without walls, where everything gathered from the huge distances and varieties of Oriental culture became categorically Oriental. It would be reconverted, restructured from the bundle of fragments brought back piecemeal by explorers, expeditions, commissions, armies, and merchants into lexicographical, bibliographical, departmentalized and textualized Orientalist sense.
Edward Said, Orientalism
There is an importance that western civilizations give to language, to nomenclature. In Madama Butterfly, there is a repeated idea that in the West butterflies are trapped and preserved for their beauty. Apart from its significance to the short story and the opera as a metaphor, this is largerly representative of the Occident’s clinical fascination with the land and creatures of the Orient, one that arguably has been extended to their treatment of people as well by not regarding them as humans but as lesser. “As Europe’s fascination with its exotic and mysterious ‘Other’ developed, particularly in the popular consciousness, it also came to be increasingly informed by the contemporary interest in the emerging sciences of botany, anthropology, comparative anatomy, palaeontology and archeology (17).” The Other becomes “exotic,” “fascinating,” the words that are so commonly extended to people of color as well. In Chinua Achebe’s Things Fall Apart, Mpalive-Hangson Msiska and David Whittaker go on to point out that “What Achebe so vividly succeeds in highlighting in [his ending] is the difference between the ways the uncomprehending European colonizers recorded the history of the colonial encounter in Africa, together with their documentation of the peoples and cultures colonized, and the reality of this historical encounter as it was by those being colonized. The final paragraph of the novel also describes the overthrow of an oral culture by a literate one (15).” Because there is an imbalance in power and knowledge, as highlighted by the relationship between Prospero and Caliban in The Tempest. There is also an imbalance in how logic is treated. As Edward Said discusses in his Introduction to Orientalism, Oriental knowledge is treated as biased, subjective and emotionally tainted while the Occident’s bias is not acknowledged in the same way, but rather treated as objective knowledge. The Occident does not really consider the Orient’s knowledge to be valuable--there is a discussion in Chinua Achebe’s novel Things Fall Apart between Akunna and Mr. Brown, in which Akunna listens and has a genuine conversation and asks questions while Mr. Brown only listens to refute points rather than treating what Akunna says seriously.
0 notes
Text
Hypsognathus fenneri

By Ashley Patch
Etymology: High Jaw
First Described By: Gilmore, 1928
Classification: Biota, Archaea, Proteoarchaeota, Asgardarchaeota, Eukaryota, Neokaryota, Scotokaryota, Opimoda, Podiata, Amorphea, Obazoa, Opisthokonta, Holozoa, Filozoa, Choanozoa, Animalia, Eumetazoa, Parahoxozoa, Bilateria, Nephrozoa, Deuterostomia, Chordata, Olfactores, Vertebrata, Craniata, Gnathostomata, Eugnathostomata, Osteichthyes, Sarcopterygii, Rhipidistia, Tetrapodomorpha, Eotetrapodiformes, Elpistostegalia, Stegocephalia, Tetrapoda, Reptiliomorpha, Amniota, Sauropsida, Diapsida?, Parareptillia, Procolophonomorpha, Ankyramorpha, Procolophonia, Procolophonoidea, Procolophonidae, Leptopleuroninae, Leptopleuronini
Time and Place: Between 228 and 202 million years ago, from the Carnian to the Rhaetian of the Late Triassic
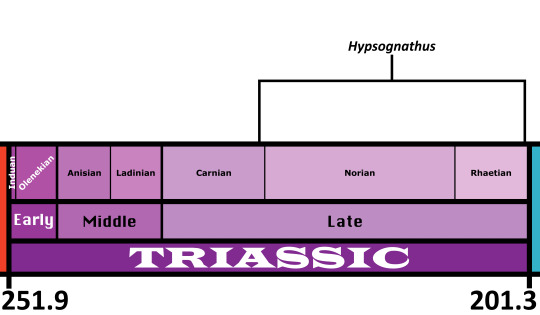
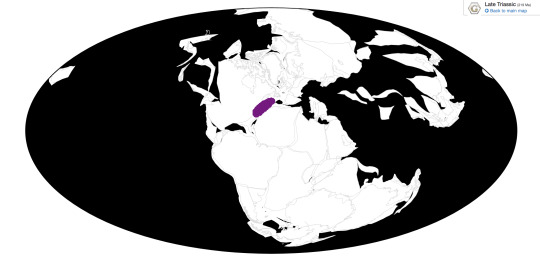
Hypsognathus is known from a wide variety of formations, including the Wolfville and Blomidon Formations of Nova Scotia, the New Haven Formation of Connecticut, and the Passaic Formation of New Jersey.
Physical Description: Hypsognathus was a Parareptile, a group of odd creatures much more common in the Permian but had their last hurrah during the Triassic. These animals were some of the most varied and fascinating creatures of their time, including some of the first bipeds, first megafauna, and first aquatic reptiles. In the Triassic, most looked like your generic tetrapod - lizards, except without their specializations and long tails; salamanders but with scales. Some, however, kept doing weird things, including our friend Hypsognathus here. Hypsognathus was about 33 centimeters long, with stocky limbs and a thick trunk. Its head was almost half as wide as the body was long at 12.5 centimeters, and it wasn’t very mobile (or kinetic) - instead, fixed in place for extra support and sturdiness. Notably, Hypsognathus had extensive spikes and protrusions coming off of its head to the sides and down on the face, making it look rather monstrous from the front. These spikes may have even been longer than the fossil indicates, covered by keratin for display purposes. Its jaw was curved upwards, giving it a weird sort of permanent smile, and it had giant teeth protruding from its mouth. These teeth were blunt and thick, allowing for strong mashing of food. These teeth were also fascinating because there was clear tooth replacement, usually alternating - in the sequence of ABABABAB, the A’s would get replaced, and then the B’s. The rest of its body was fairly standard for a Procolophonid - with short, splayed out legs for walking slowly and from side to side; wide and thick fingers and toes for gripping the ground; and a short stubby tail not used for much at all. It would have probably been covered in something akin to scales, and well adapted for dry conditions as a result.
Diet: Hypsognathus was an herbivore, feeding on high-fiber, tough plant material.
Behavior: Hypsognathus was, more likely than not, a burrowing animal. The lack of kinesis in the skull allowed it to use it like a shovel, which may have been one of the uses of the spikes on its face. It could then dig into the ground to hide from predators, burrowing deep and not worrying about the fact that the rest of its body is relatively unprotected since it is being hidden by the dirt. This also explains the lack of ornamentation elsewhere on the body, and its squat and short structure. A long tail, or long limbs, would not have aided in hiding in the dirt! In addition, those wide and thick fingers and toes would have helped in kicking up dirt and escaping from predators quickly. The spikes may have also been able to anchor Hypsognathus within the burrow itself, preventing it from being dug out by a small predator. These spikes would have also been decent as display structures, with longer or more ridiculous looking ones appearing Fancy to other Hypsognathus. This could have been added on to with more keratin sheaths, reflecting the ability of an individual Hypsognathus to waste energy - and burrow space - on more elaborate horns because it was doing so well. It would then emerge from the burrows to feed on roughage and tough plants - though it may have been able to feed on roots and tubers underground as well. Given it could have used the horns for display, it was probably at least somewhat social; however, we have no idea how much or if it took care of its young, or had any other complex behaviors. Juvenile and young specimens are known, and they also have spikes, so if they served for communication, social life may have been a part of youth as well as adulthood.
Ecosystem: Hypsognathus was a consistent feature of Northeastern North America during the Late Triassic, present in a variety of environments and ecosystems along the geologically active area that would eventually open up to begin forming the Atlantic Ocean. It generally favored sandy beaches lining seasonal lakes and rivers, with a variety of coniferous trees and swamp trees rooted in the lake. There were also ferns, cycads, and plankton abundant around and within the water. There were also proto angiosperms! While Hypsognathus lived with a wide variety of animals, some creatures kept popping up over and over again - the Aetosaur Stegomus, Phytosaurs such as Belodon and Rutiodon, Rhynchosaurs like Scaphonyx and Colobops, predatory Pseudosuchians such as Erpetosuchus and Rauisuchians, the weirdo Tanystropheid Gwyneddichnium and the Allokotosaur Teraterpeton, other Procolophonids like Scoloparia and Acadiella, the Temnospondyl Metoposaurus, the Cynodont Arctotraversodon, and there were also early dinosaurs and late Dicynodonts (though only footprints were preserved). In fact, Hypsognathus is most commonly preserved alongside footprints, lending credence to the idea that it would burrow in wet, sandy places, and get trapped and preserved there in the same events that preserved the prints.
Other: This strange burrower with a spikey head was a very successful animal - it was found all over Eastern North America, throughout the Late Triassic, and seems to have only gone extinct because of the end-Triassic extinction. This is notable, because it is one of the last surviving Parareptiles, ever. The other Parareptiles of the Triassic were also small, squat lizard-like burrowers, many with interesting head ornamentation as well. While the heyday of Parareptiles was behind them, they managed to put on an excellent (and adorable) final act. In fact, morphological diversity of Parareptiles went down from a wide variety of shapes and forms and lifestyles to just one, the Procolophonids like Hypsognathus. This makes Hypsognathus a unique example of an ancient lineage, survivors of one Mass Extinction just to be finished off by another.
~ By Meig Dickson
Sources Under the Cut
Carroll, R. L., E. S. Belt, D. L. Dineley, D. Baird, and D. C. McGregor. 1972. In D. J. Glass (ed.), Guidebook: Excursion A59. Vertebrate Palaeontology of Eastern Canada 1-113.
Colbert, E. H. 1946. Hypsognathus, a Triassic reptile from New Jersey. Bulletin of the American Museum of Natural History 86: 225 - 274.
Ford, D. P., R. B. J. Benson. 2020. The phylogeny of early amniotes and the affinities of Parareptilia and varanopidae. Nature Ecology & Evolution 4: 57 - 65.
Gilmore, C. W. 1928. New Fossil Reptile from the Triassic of New Jersey. Proceedings of the United States National Museum 73(7):1-8.
Lucas, S. G. 2018. Late Triassic Terrestrial Tetrapods: Biostratigraphy, Biochronology, and Biotic Events. The Late Triassic World, Topics in Geobiology 46: 351 - 405.
Macdougall, M. J.; D. Scott, S. P. Modesto, S. A. Williams, R. R. Reisz. 2017. New material of the reptile Colobomycter pholeter (Parareptilia: Lanthanosuchoidea) and the diversity of reptiles during the Early Permian (Cisuralian). Zoological Journal of the Linnean Society 20: 1 - 11.
MacDougall, M. J.; N. Brocklehurst, J. Frobisch. 2019. Species richness and disparity of pararpetiles across the end-Permian Mass Extinction. Proceedings of the Royal Society B 286: 2018572.
Olsen, P. E., and D. Baird. 1986. The ichnogenus Atreipus and its significance for Triassic biostratigraphy. In: K. Padian (ed.), The Beginning of the Age of Dinosaurs: Faunal Changes Across the Triassic-Jurassic Boundary. Cambridge University Press, Cambridge 61-87.
Olson, P. E. 1988. Paleoecology and Paleoenvironments of the Continental Early Mesozoic Newark Supergroup of Eastern North America: In Manspeizer, W. (ed.), Triassic-Jurassic Rifting and the Opening of the Atlantic Ocean, Elsevier, Amsterdam, p. 185-230.
Palmer, D. 1999. The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals. London: Marshall Editions.
Sues, H. D., P. E. Olsen, D. M. Scott, P. S. Spencer. 2000. Cranial Osteology of Hypsognathus fenneri, a latest Triassic procolophonid reptile from the Newark Supergroup of Eastern North America. Journal of Vertebrate Paleontology 20 (2): 275 - 284.
Sues, H.D. 2019. The Rise of Reptiles: 320 Million Years of Evolution. JHU Press.
Tsuji, L. A. 2017. Mandaphon nadra, gen. Et sp. Nov., a new procolophonid from the Manda Beds of Tanzania. Journal of Vertebrate Paleontology 37 (supp1): 80 -87.
Zaher, M., R. A. Coram, M. J. Benton. 2018. The Middle Triassic procolophonid Kapes bentoni: computed tomography of the skull and skeleton. Papers in Palaeontology 5 (1): 111 - 138.
#hypsognathus#parareptile#procolophonid#triassic#triassic madness#triassic march madness#prehistoric life#paleontology
112 notes
·
View notes
Text
Foro panarium

By Ripley Cook
Etymology: Breadbasket’s Hole
First Described By: Olson, 1992
Classification: Dinosauromorpha, Dinosauriformes, Dracohors, Dinosauria, Saurischia, Eusaurischia, Theropoda, Neotheropoda, Averostra, Tetanurae, Orionides, Avetheropoda, Coelurosauria, Tyrannoraptora, Maniraptoromorpha, Maniraptoriformes, Maniraptora, Pennaraptora, Paraves, Eumaniraptora, Averaptora, Avialae, Euavialae, Avebrevicauda, Pygostaylia, Ornithothoraces, Euornithes, Ornithuromorpha, Ornithurae, Neornithes, Neognathae, Neoaves, Columbaves, Otidimorphae, Musophagiformes
Status: Extinct
Time and Place: Between 53.5 and 48.5 million years ago, in the Ypresian of the Eocene

Foro is known from the Fossil Butte Member of the Green River Formation of Wyoming
Physical Description: Foro had a downward sloping, distinctive skull, with a slight point to the tip at the end of the beak, hanging off of a long neck. It had a large, rounded head, a fairly stout body, and long spindly legs. Its wings were decently sized too, making this in general a very lanky sort of bird. However, these wings were probably weak, and in addition to the stocky body it wouldn’t have been the world’s greatest flier. As for its general size, it was probably about the size of a modern roadrunner, reaching between 22 and 24 centimeters in length; it was probably similar in many ways to living roadrunners, while simultaneously being like a bustard, a hoatzin, and a relative of the modern day turacos. Evolution is wild, man.
Diet: Given it had the general head-shape of a Hoatzin, it seems at least somewhat likely that Foro was herbivorous, though this was of course evolved convergently to the Hoatzin (as Foro was probably a stem-turaco). This is further corroborated by the fact that turacos are also mostly herbivorous.
Behavior: Foro was a ground-dwelling bird, rather than tree-dwelling like modern turacos; it would use its long legs to wander about its lake environment, searching for food to eat and moving through more difficult ground material such as mud and dirt. In this way it was much more like a living bustard than a living turaco, indicating the bustard way of life was actually the ancestral case for cuckoos and turacos before they adapted for their unique arboreal roles. As a weird cross between a bustard and a wading bird, it would have spent a good amount of time moving about the ground, grabbing food from the mud and in the low-lying plants, and running away rapidly at the first sign of danger. It was still able to fly, and would use that when the danger was way too close. As a bird, it would have been an active animal; it is especially uncertain whether or not it was particularly social, but it probably took care of its young.

By Scott Reid
Ecosystem: The Green River Formation is a famous environment from the early Eocene of North America, showcasing the burst of creatures after the end-Cretaceous extinction that came forward as the global rainforest reached its peak, and serving as a host of creatures that were precursors to modern forms. This was a fossil lake system scattered across the jungle in loosely dense vegetation, including sycamore trees and ferns, palms, and many other kinds of plants. As the Rocky Mountains formed nearby, nutrients would randomly spike in the lake, dumping phosphorus and suffocating the fish inside. This lead to unique preservation and an amazing number of fossils from Green River. Here there were a variety of rays, catfish, herrings, and countless other fish; crocodilians like Borealosuchus; early primates, bats, and an armadillo-esque mammal. Birds were also extremely common: there was the gull-like frigatebird Limnofregata, the Lithornithids Pseudocrypturus and Calciavis, the flamingo-duck Presbyornis, the pre-pheasant Gallinuloides, the early swift-hummingbird Eocypselus, the frogmouth-esque Fluvioviridavis and Prefica, the mousebird Anneavis, the woodpecker Neanis, the Parrots of Prey Cyrilavis and Tynskya, and the parrot-footed passerines Zygodactylus and Eozygodactylus. In this way, Foro was a notable and usual member of the ecosystem, showcasing the weird mode of evolution of the modern Turaco group.
Other: Foro has been the subject of a fascinating research history, with it originally being called its own thing in the Cuckoos, and then thought to be a Hoatzin for a while, and then finally assigned as a stem-turaco. This is of utter importance because it showcases how neotropical birds like the turacos (which, today, are limited to Africa) were once widespread - this also applies to many other groups of birds, and this collapse of tropical birds in North America and Europe is an object of intense study. As the Global rainforest collapsed, the evolutionary trajectory of modern birds changed dramatically, and Foro was a part of that. On the other hand, Foro and other birds known from the Paleocene and Eocene epochs showcase how dinosaurs managed to bounce back from the extinction; birds were more ground-dwelling than tree-dwelling at first, given the lack of trees in impact winter; Foro was a ground-dwelling bird, but one on the path to tree-life, much like many other dinosaurs in this early part of the new era.
~ By Meig Dickson
Sources under the Cut
Felice, R. N. 2014. Coevolution of caudal skeleton and tail feathers in birds. Journal of Morphology 275 (12): 1-10.
Field, D. J., A. Bercovici, J. S. Berv, R. Dunn, D. E. Fastovsky, T. R. Lyson, V. Vajda, J. A. Gauthier. 2018. Early Evolution of Modern Birds Structured by Global Forest Collapse at the End-Cretaceous Mass Extinction. Current Biology 28: 1825 - 1831.
Field, D. J., A. Y. Hsiang. 2018. A North American stem turaco, and the complex biogeographic history of modern birds. BMC Evolutionary BIology 18: 102.
Grande, L., P. Bucheim. 1994. Palaeontological and Sedimentological Variation in Early Eocene Fossil Lake. Contributions to Geology, University of Wyoming 30: 45.
Hutchison, J. H. 2013. New turtles from the Paleogene of North America. In D. B. Brinkman, P. A. Holroyd, J. D. Gardner (eds.), Morphology and Evolution of Turtles 477-497.
Mayr, G. 2009. Paleogene Fossil Birds. Springer-Verlag Berlin Heidelberg.
Mayr, G. 2017. Avian Evolution: The Fossil Record of Birds and its Paleobiological Significance. Topics in Paleobiology, Wiley Blackwell. West Sussex.
Miller, A. H. 1953. A fossilized hoatzin from the Miocene of columbia. Auk 70: 484 - 489.
Olson, S. L. 1977. A Lower Eocene frigatebird from the Green River Formation of Wyoming (Pelecaniformes: Fregatidae). Smithsonian Contributions to Paleobiology 35:1-33.
Olson, S. L. 1992. A new family of primitive landbirds from the Lower Eocene Green River Formation of Wyoming. Papers in avian paleontology honoring Pierce Brodkorb: 127 - 136.
Olson, S. L., H. Matsuoka. 2005. New specimens of the early Eocene frigatebird Limnofregata (Pelecaniformes: Fregatidae), with the description of a new species. Zootaxa 1046: 1 - 15.
Smith, M. E., B. Singer, A. Carroll. 2003. 40Ar/39Ar Geochronology of the Eocene Green River Formation, Wyoming. Geological Society of America Bulletin 115 (5): 549 - 565. Stidham, T. A. 2014. A new species of Limnofregata (Pelecaniformes: Fregatidae) from the Early Eocene Wasatch Formation of Wyoming: implications for palaeoecology and palaeobiology. Palaeontology: 1 - 11.
#Foro panarium#Foro#Bird#Turaco#Dinosaur#Birds#Dinosaurs#Palaeoblr#Birblr#Factfile#Columbavian#Paleogene#North America#Terrestrial Tuesday#paleontology#prehistory#prehistoric life#biology#a dinosaur a day#a-dinosaur-a-day#dinosaur of the day#dinosaur-of-the-day#science#nature
142 notes
·
View notes
Text
FossilWatch: Stats, sex, skulls and sleep
Sleeping through tough times
US scientists have discovered fossil evidence of a hibernation-like state in an animal that lived in Antarctica during the Early Triassic, some 250 million years ago.
It is the oldest such find, they say, and suggests that torpor – a general term for hibernation and similar states in which animals temporarily lower their metabolic rate to get through a tough season – arose in vertebrates even before mammals and dinosaurs evolved.
The creature, a member of the genus Lystrosaurus, was a distant relative of mammals.
It spread across Earth’s then-single continent, Pangea, and fossils have been found in India, China, Russia and Africa, as well as Antarctica, which at the time experienced extended periods without sunlight each winter.
Life restoration of Lystrosaurus in a state of torpor. Credit: Crystal Shin
“Animals that live at or near the poles have always had to cope with the more extreme environments present there,” says lead author Megan Whitney. “These preliminary findings indicate that entering into a hibernation-like state is not a relatively new type of adaptation. It is an ancient one.”
Whitney carried out the work while at the University of Washington, with then colleague Christian Sidor. Their findings are published in the journal Communications Biology.
Lystrosaurus had no teeth but bore a pair of tusks in the upper jaw, Witney says, which made their study possible. As in elephants, the tusks grew continuously, so cross-sections of fossilised tusks provide information about metabolism, growth and stress or strain.
The researchers compared tusks from six Antarctic Lystrosaurus and four from South Africa and noticed similar growth patterns but one key difference: closely-spaced, thick rings that likely indicate periods of less deposition due to prolonged stress.
“The closest analogue we can find to the ‘stress marks’ that we observed… are stress marks in teeth associated with hibernation in certain modern animals,” Whitney says.
The face of a dinosaur embryo
The first dinosaur embryos were found in a nesting ground of titanosaurian at Auca Mahuevo in Argentinian Patagonia. Two decades later, an international research team has described the first near-intact embryonic skull.
A magnified perspective of the Titanosaurian embryonic skull and a skull reconstruction. Credit: Kundrat et al
The findings, published in the journal Current Biology, suggest that sauropod dinosaurs, a group characterised by the long neck and tails and small heads familiar in the Brontosaurus, may have had specialised facial features as hatchlings that changed as they grew into adults.
“The most striking feature is head appearance, which implies that hatchlings of giant dinosaurs may differ in where and how they lived in their earliest stages of life,” says lead author Martin Kundrat of Pavol Jozef Šafárik University, Slovak Republic.
“But because it differs in facial anatomy and size from the sauropod embryos of Auca Mahuevo, we cannot rule out that it may represent a new titanosaurian dinosaur.”
Kundrat’s team used an imaging technology called synchrotron microtomography to study the inner structure of bones, teeth and soft tissues of the embryonic dinosaur and found hidden details, including tiny teeth preserved deeply in tiny jaw sockets.
The specimen also is from Patagonia, although its precise origin isn’t known. It was illegally exported and came to researchers’ attention only later. It’s now housed with other embryos from Auca Mahuevo at Argentina’s Museo Municipal Carmen Funes.
Thinking about dinosaur sex
Speaking of dinosaurs, how do you tell boys from girls? The answer may be to use stats.
In a new paper in the Biological Journal of the Linnean Society, scientists led by the Field Museum, US, explain how a different kind of analysis can estimate the degree of sexual variation in a dataset of fossils.
“It’s a whole new way of looking at fossils and judging the likelihood that the traits we see correlate with sex,” says lead author Evan Saitta. “This paper is part of a larger revolution of sorts about how to use statistics in science but applied in the context of palaeontology.”
Palaeontologists have previously used significance testing, where you collect all your data points then calculate the probability that those results could have happened by pure chance rather than an actual cause. This sometimes works for big, clean datasets, Saitta says, but “with a lot of these dinosaur tests, our data is pretty bad”.
He and his colleagues turned to effect size statistics, which attempts to estimate the degree of sex differences and calculate the uncertainty in that estimate. This approach takes natural variations into account without viewing dimorphism as black-or-white. Many sexual dimorphisms can be subtle.
“We showed that if you adopt this paradigm shift in statistics, where you attempt to estimate the magnitude of an effect and then put error bars around that, you can often produce a fairly accurate estimate of sexual variation even when the sexes of the individuals are unknown,” says Saitta. The work continues.
Old fossil adds to old debate
US and Canadian researchers have uncovered fossils of a new species of marine animal, Gyaltsenglossus senis, that could help solve a longstanding debate.
Credit: Jean-Bernard Caron. Royal Ontario Museum
Discovered at a Burgess Shale site in the Canadian Rockies, they are over 500 million years old and just two centimetres in length, but the remarkably preserved soft tissues reveal incredibly detailed anatomical structures.
These details include the oval-shaped proboscis of acorn worms (aka enteropneusta) and a basket of feeding tentacles similar to those of pterobranchs. And that’s where things get interesting, because this provides evidence connecting these two major groups of hemichordates.
“Acorn worms and pterobranchs look so different from each other that understanding the origins of their evolutionary relationship has been a major historical question in zoology,” says Karma Nanglu from the Smithsonian National Museum of Natural History, lead author of a paper in the journal Current Biology.
“Answering this question has been made much harder by the extreme lack of fossils of these soft-bodied hemichordates. Throughout the half-billion-year-long history of hemichordates you can count on one hand the number of exceptional preserved fossil species.”
An ancient animal with an intermediary anatomy between acorn worms and pterobranchs had been hypothesised before but “this new animal is the clearest view of what the ancestral hemichordate may have looked like,” says co-author Christopher Cameron, from the University of Montreal.
Hemichordates belong to a major division of animal life called Deuterostomia, which includes chordates like fish and mammals. When looking at Gyaltsenglossus, we’re actually looking at a very, very distant relative of our own branch of vertebrate and human evolution, Nanglu says.
FossilWatch: Stats, sex, skulls and sleep published first on https://triviaqaweb.weebly.com/
0 notes
Last Seen Blogs

jaymelikes-blog
JAYME LIKES...

k-writer17
Hiddlestoner1615

imaginingtomhardy
Tom Hardy fanfiction

jetaime-jespere
Hotchniss&CM

charleettscoolpage
COOL PAGE