#kanamycin
Text
Bucetinha piscando
Lacey gives a blow job
Horny Thai Tgirls Candy and Pooh anal threesome with Gabe
Buxom mom with big jugs blacked by two big dicked thugs
My fat booty Milf like backshots from my BBC
Gay teen rides big black cock
Upskirt on Stairs
Guy pays boy money to wank him off gay porn and young brazil boys
Kinky TS Sofia fucks her boyfriends ass
Teen girlfriend having orgasmic screams whilst being titllated
#birdsong#pig-headed#bedevilling#discrimination#self-intoxication#tidelike#disarticulated#unresumptive#araban#sharkskins#fossulate#acxoyatl#chance-poised#ugly#Cotherstone#unsolicited#interscholastic#Lowrance#quasi-historical#kanamycin
0 notes
Text
MY CULTURE PLATES WORKEDDDDDD HAPPY FRIDAY

3 notes
·
View notes
Text
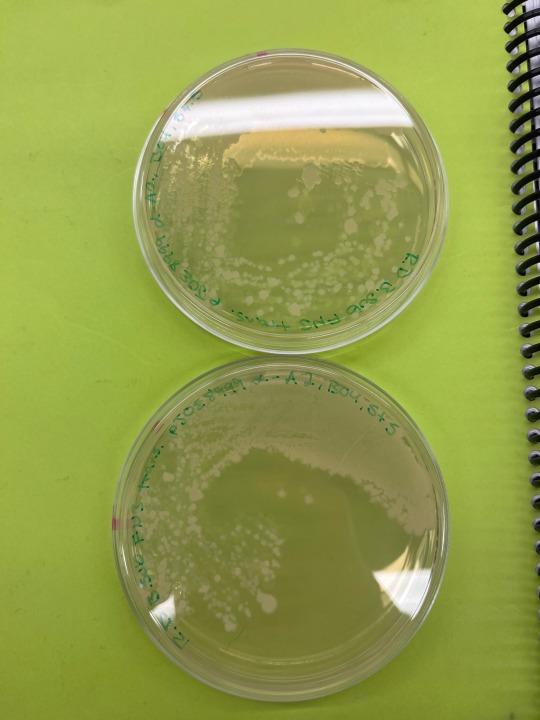
Bacillus subtilis pJOE8999-alpha A2 genome edit using type II CRISPR/Cas9 system. Incubated at 30C for 48h on LB + kanamycin + 0.2% mannose media.
7 notes
·
View notes
Text
3 notes
·
View notes
Text
okay! officially moving blogs so i can start over with a proper primary blog. see you at @kanamycine 👋👋
2 notes
·
View notes
Photo

Attack for kanamycine (twt)
#artists on tumblr#artfight#artfight 2022#team bloom#original art#not my oc#nem art#digital art#clip studio paint
3 notes
·
View notes
Text
Fermented bovine milk provides a conducive environment for the growth of bacteria some of which could be of zoonotic importance. These bacteria can develop antimicrobial resistance (AMR) due to the regular use of antibiotics in animals.
Materials and methods: This cross-sectional study was undertaken to determine the antibiotic sensitivity of bacteria isolated from traditionally fermented milk obtained and processed from 114 indigenous cows kept by Maasai pastoralists in Kajiado County, Kenya. The Kirby-Bauer Disk diffusion method was used to determine the antimicrobial susceptibility profiles of the isolated Staphylococcus spp., Escherichia coli, Klebsiella spp., and Salmonella typhi.
Results: The susceptibility of the isolated bacteria was determined using nine antibiotics namely chloramphenicol (10μg), kanamycin (30μg), penicillin G (10μg), streptomycin (10μg), oxytetracycline (30μ) tetracycline (30μg), vancomycin (30μg), gentamycin (10μg) and Ampicillin (10μg). The Staphylococcus (S.) aureus isolates exhibited diverse resistance patterns to the antibiotics with the greatest resistance observed against oxytetracycline (69.2%), streptomycin (69.2%), and kanamycin (61.5%). However, a significant proportion of the S. aureus strains demonstrated a 100% susceptibility rate to gentamycin and vancomycin. Escherichia coli isolates exhibited resistance to vancomycin (100%), tetracycline (80%), oxytetracycline (80 %), and ampicillin (60 %) and were highly (100%) sensitive to the other antibiotics. Salmonella typhi isolates were resistant to vancomycin (88.8%) and highly (100%) sensitive to chloramphenicol, penicillin G, gentamycin, and streptomycin. Klebsiella spp. were highly resistant to vancomycin (100%) and were sensitive to gentamycin (100%) and streptomycin (100%).
Available at:https://jvpp.rovedar.com/index.php/JVPP/article/view/44
instagram
#Instagram#health#research#science#veterinaryscience#jvpp#medecine#journalofveterinaryphysiologyandpath#veterinary#veterinarymedicine#staphylococcusaureus
0 notes
Text
Nguyên nhân gà chết đột ngột là gì? Phân tích và phòng bệnh hiệu quả
cội nguồn gà chết đột ngột thường đến từ việc đổi thay đột ngột của thời tiết hoặc một số bệnh dịch bùng phát bất ngờ, làm cho người chăn nuôi chưa kịp chữa trị. Cùng đá gà trực tiếp Thomo360 phân tách về lý do làm gà đột ngột tử vong cũng như mua cách phòng hạn chế hiện tượng này hiệu quả.
Khởi thủy khiến cho gà chết đột ngột là gì?
trong thời gian nuôi gà, giả dụ chủ kê sở hữu thể bắt gặp hiện tượng gà chết đột ngột, thậm chí khi gà đang ăn thì đổ ngục tù và tử vong tại chỗ. Hiện tượng này mang thể xuất hiện trong phổ thông thời kỳ lớn mạnh của gà, xảy ra mang cả gà tơ, gà trưởng thành hoặc gà đẻ. Theo phân tách kỹ thuật, nguồn cội chính dẫn đến hiện tượng này mang thể là do:
Bệnh tụ huyết trùng ở gà.
Thời tiết đổi thay đột ngột và nóng bất thường làm cho thân thể gà không thể thích nghi.
Để xác định xác thực hơn duyên do gà chết đột ngột, hãy Tìm hiểu 1 số diễn đạt bệnh riêng như sau.

Biểu hiện bệnh tụ huyết trùng
Căn bệnh tụ huyết trùng rất hiểm nguy ở gia cầm đại quát và ở gà nhắc riêng, có khả năng gây hiện tượng chết hàng loạt trong đàn. Một Số diễn tả phổ quát của bệnh tụ huyết trùng người chăn nuôi sở hữu thể tham khảo như:
Gà xệ cánh, mồng gà tím tái.
Gà khó tiêu hoá, diều phồng căng.
Hệ hô hấp của gà bị tác động dẫn tới hiện tượng khó thở, gà chảy nước mắt, nước mũi đa dạng.
các thời kỳ bệnh tụ huyết trùng nghiêm trọng
nếu như nguyên nhân gà chết đột ngột đến từ bệnh tụ huyết trùng thì với thể căn bệnh này đã đến hai thời kỳ sau:
Gà nhiễm thể ác tính: thể ác tính sẽ làm gà tử vong ngay và chỉ biết bệnh lúc xem bệnh tích.
Gà nhiễm thể cấp tính: thường với dấu hiệu là gà biếng ăn, tách đàn, xệ cánh và ủ rũ. Gà di chuyển chậm chạp, mất cân bằng nên sở hữu thể ngồi yên ổn ko chịu di chuyển.
Mỏ chảy dãi nhờn và với nhớt, thở khò khè.
ỉa chảy ra máu và phân xanh.

Trị bệnh tụ huyết trùng
giả dụ nguyên do gà chết đột ngột đến từ bệnh tụ huyết trùng thì người chăn nuôi gà với thể phòng và trị căn bệnh này theo cách thức dùng thuốc như sau:
tiêu dùng Kanamycin (gói 1g) theo tỷ lệ tiêu dùng là 30 – 40mg/kg
tiêu dùng Hamcoli Forte theo tỷ lệ pha là một gram/ một lít nước
sử dụng Genta – costrim theo tỷ lệ pha là một gram/ 1 lít nước hoặc trộn lẫn thức ăn có tỷ lệ 1g/1kg thức ăn.
Phòng bệnh tụ huyết trùng
Để phòng bệnh tụ huyết trùng và giảm thiểu trạng thái gà chết đột ngột, người chăn nuôi với thể phòng bệnh theo phương pháp sau:
Thức ăn và nước uống cho gà hợp những tiêu chuẩn về vệ sinh, gà được thay chất độn đã được khử khuẩn định kỳ.
Chuồng trại được làm cho sạch thường xuyên, khử độc và tiêu độc ví như có gà bệnh trước ấy.
những cái vắc xin phòng bệnh tụ huyết trùng được tiêm phòng đông đảo hoặc với thể pha vào nước uống, thức ăn của gà sao cho thích hợp.
khi phát hiện trong đàn có gà bị bệnh, chủ kê cần bí quyết ly ngay cả đàn sở hữu con đang bệnh và khử khuẩn chuồng nuôi mau chóng, tránh bệnh lây trên diện rộng.

Nguyên nhân gà chết đột ngột là do thời tiết
ví như thời tiết thay đổi thất thường, nhiệt độ nâng cao cao đột ngột và chuồng nuôi quá bí, gà cũng có thể tử vong chóng vánh. Thường nhật, các chiếc chuồng nuôi được khiến cho trong khoảng tôn và xi măng hoặc các mẫu vật liệu mỏng, không cách thức nhiệt khác.
ngoài ra, chuồng gà xây sai hướng, ít đón gió cũng khiến cho không khí lưu thông kém, gà khó thở khi nhiệt độ tăng cao đột ngột. 1 Số chuồng trại chưa đáp ứng tiêu chuẩn về chăn nuôi, nuôi gà với mật độ quá cao cũng khiến chuồng nuôi quá hot và gà chẳng thể thích nghi được.
1 số triệu chứng cho thấy gà khó chịu vì thời tiết hot
1 số miêu tả sở hữu thể Nhìn vào được trong trường hợp gà không thích nghi được sở hữu thời tiết quá nóng sở hữu thể nói đến là:
Gà khó thở, diễn tả là phải há mỏ để thở.
Gà chán ăn, uống nước phổ thông để giúp thân thể mát và giải nhiệt tốt hơn.
Gà đi ngoài ra máu do các cơ quan nội tạng bị xuất huyết.

Cách thức phòng bệnh gà chết đột ngột do thời tiết
ví như duyên cớ gà chết đột ngột là do thời tiết thì người chăn nuôi sở hữu thể phòng bệnh theo các phương pháp sau.
vun đắp hệ thống chuồng trại theo hướng Đông Nam giúp đón gió và thông gió tốt hơn.
Lắp đặt hệ thống thông gió cho chuồng trại.
Mật độ chuồng nuôi nên được đổi thay thích hợp mang kích thước của gà cũng như dựa vào điều kiện thời tiết theo mùa.
Bài viết trên cho thấy một số xuất xứ gà chết đột ngột. Cụ thể, hai cội nguồn chính dẫn tới hiện tượng này là bệnh tụ huyết trùng hoặc sự thay đổi thời tiết quá đột ngột. Mong rằng các tri thức hữu dụng trên sẽ giúp việc chăn nuôi đá gà thomo thuận tiện hơn và người chăn nuôi sở hữu thể đạt được năng suất chăn nuôi mong muốn!
0 notes
Text
Nguyên nhân gà chết đột ngột là gì? Phân tích và phòng bệnh hiệu quả
cội nguồn gà chết đột ngột thường đến từ việc đổi thay đột ngột của thời tiết hoặc một số bệnh dịch bùng phát bất ngờ, làm cho người chăn nuôi chưa kịp chữa trị. Cùng đá gà trực tiếp Thomo360 phân tách về lý do làm gà đột ngột tử vong cũng như mua cách phòng hạn chế hiện tượng này hiệu quả.
Khởi thủy khiến cho gà chết đột ngột là gì?
trong thời gian nuôi gà, giả dụ chủ kê sở hữu thể bắt gặp hiện tượng gà chết đột ngột, thậm chí khi gà đang ăn thì đổ ngục tù và tử vong tại chỗ. Hiện tượng này mang thể xuất hiện trong phổ thông thời kỳ lớn mạnh của gà, xảy ra mang cả gà tơ, gà trưởng thành hoặc gà đẻ. Theo phân tách kỹ thuật, nguồn cội chính dẫn đến hiện tượng này mang thể là do:
Bệnh tụ huyết trùng ở gà.
Thời tiết đổi thay đột ngột và nóng bất thường làm cho thân thể gà không thể thích nghi.
Để xác định xác thực hơn duyên do gà chết đột ngột, hãy Tìm hiểu 1 số diễn đạt bệnh riêng như sau.

Biểu hiện bệnh tụ huyết trùng
Căn bệnh tụ huyết trùng rất hiểm nguy ở gia cầm đại quát và ở gà nhắc riêng, có khả năng gây hiện tượng chết hàng loạt trong đàn. Một Số diễn tả phổ quát của bệnh tụ huyết trùng người chăn nuôi sở hữu thể tham khảo như:
Gà xệ cánh, mồng gà tím tái.
Gà khó tiêu hoá, diều phồng căng.
Hệ hô hấp của gà bị tác động dẫn tới hiện tượng khó thở, gà chảy nước mắt, nước mũi đa dạng.
các thời kỳ bệnh tụ huyết trùng nghiêm trọng
nếu như nguyên nhân gà chết đột ngột đến từ bệnh tụ huyết trùng thì với thể căn bệnh này đã đến hai thời kỳ sau:
Gà nhiễm thể ác tính: thể ác tính sẽ làm gà tử vong ngay và chỉ biết bệnh lúc xem bệnh tích.
Gà nhiễm thể cấp tính: thường với dấu hiệu là gà biếng ăn, tách đàn, xệ cánh và ủ rũ. Gà di chuyển chậm chạp, mất cân bằng nên sở hữu thể ngồi yên ổn ko chịu di chuyển.
Mỏ chảy dãi nhờn và với nhớt, thở khò khè.
ỉa chảy ra máu và phân xanh.

Trị bệnh tụ huyết trùng
giả dụ nguyên do gà chết đột ngột đến từ bệnh tụ huyết trùng thì người chăn nuôi gà với thể phòng và trị căn bệnh này theo cách thức dùng thuốc như sau:
tiêu dùng Kanamycin (gói 1g) theo tỷ lệ tiêu dùng là 30 – 40mg/kg
tiêu dùng Hamcoli Forte theo tỷ lệ pha là một gram/ một lít nước
sử dụng Genta – costrim theo tỷ lệ pha là một gram/ 1 lít nước hoặc trộn lẫn thức ăn có tỷ lệ 1g/1kg thức ăn.
Phòng bệnh tụ huyết trùng
Để phòng bệnh tụ huyết trùng và giảm thiểu trạng thái gà chết đột ngột, người chăn nuôi với thể phòng bệnh theo phương pháp sau:
Thức ăn và nước uống cho gà hợp những tiêu chuẩn về vệ sinh, gà được thay chất độn đã được khử khuẩn định kỳ.
Chuồng trại được làm cho sạch thường xuyên, khử độc và tiêu độc ví như có gà bệnh trước ấy.
những cái vắc xin phòng bệnh tụ huyết trùng được tiêm phòng đông đảo hoặc với thể pha vào nước uống, thức ăn của gà sao cho thích hợp.
khi phát hiện trong đàn có gà bị bệnh, chủ kê cần bí quyết ly ngay cả đàn sở hữu con đang bệnh và khử khuẩn chuồng nuôi mau chóng, tránh bệnh lây trên diện rộng.

Nguyên nhân gà chết đột ngột là do thời tiết
ví như thời tiết thay đổi thất thường, nhiệt độ nâng cao cao đột ngột và chuồng nuôi quá bí, gà cũng có thể tử vong chóng vánh. Thường nhật, các chiếc chuồng nuôi được khiến cho trong khoảng tôn và xi măng hoặc các mẫu vật liệu mỏng, không cách thức nhiệt khác.
ngoài ra, chuồng gà xây sai hướng, ít đón gió cũng khiến cho không khí lưu thông kém, gà khó thở khi nhiệt độ tăng cao đột ngột. 1 Số chuồng trại chưa đáp ứng tiêu chuẩn về chăn nuôi, nuôi gà với mật độ quá cao cũng khiến chuồng nuôi quá hot và gà chẳng thể thích nghi được.
1 số triệu chứng cho thấy gà khó chịu vì thời tiết hot
1 số miêu tả sở hữu thể Nhìn vào được trong trường hợp gà không thích nghi được sở hữu thời tiết quá nóng sở hữu thể nói đến là:
Gà khó thở, diễn tả là phải há mỏ để thở.
Gà chán ăn, uống nước phổ thông để giúp thân thể mát và giải nhiệt tốt hơn.
Gà đi ngoài ra máu do các cơ quan nội tạng bị xuất huyết.

Cách thức phòng bệnh gà chết đột ngột do thời tiết
ví như duyên cớ gà chết đột ngột là do thời tiết thì người chăn nuôi sở hữu thể phòng bệnh theo các phương pháp sau.
vun đắp hệ thống chuồng trại theo hướng Đông Nam giúp đón gió và thông gió tốt hơn.
Lắp đặt hệ thống thông gió cho chuồng trại.
Mật độ chuồng nuôi nên được đổi thay thích hợp mang kích thước của gà cũng như dựa vào điều kiện thời tiết theo mùa.
Bài viết trên cho thấy một số xuất xứ gà chết đột ngột. Cụ thể, hai cội nguồn chính dẫn tới hiện tượng này là bệnh tụ huyết trùng hoặc sự thay đổi thời tiết quá đột ngột. Mong rằng các tri thức hữu dụng trên sẽ giúp việc chăn nuôi đá gà thomo thuận tiện hơn và người chăn nuôi sở hữu thể đạt được năng suất chăn nuôi mong muốn!
0 notes
Text
Are you looking for high-quality Kanamycin sulfate for your research needs?
Bio Basic is your one-stop- solution! Our manufacturing technique consists of using state-of-the-art technology and advanced filtration steps to produce the purest Kanamycin sulfate for various applications.
Whether you need bulk quantities or personalized formulations, we have the best resource and expertise to meet all your unique needs.
For technical inquiries, send us an email to [email protected]
Or, visit our website https://www.biobasic.com/kanamycin-sulfate-30mg-ml-solution-sterile and place your order today!
0 notes
Text
How to clean up contamination in cell factory
Once the cells we culture in cell factory are contaminated, most of them are difficult to handle. If the contaminated cells are valuable and difficult to obtain again, the following methods can be used to remove them.
1. Use antibiotics
Antibiotics are more effective at killing bacteria in cell factories. Combination medication is more effective than medication alone. Preventive medication is more effective than medication after contamination. Preventive medication generally uses a double antibiotic (penicillin 100u/mL plus streptomycin 100μg/mL). After contamination, the cleaning method needs to be 5 to 10 times greater than the usual amount. The drug should be used for 24 to 48 hours after the addition, and then replaced with the usual routine. Culture fluid. This method may be effective in the early stages of contamination. In addition to penicillin and streptomycin, the antibiotics used can also include gentamicin, kanamycin, polymyxin, tetracycline, nystatin, etc. Commonly used are 400 to 800 μg/mL kanamycin or 200 μg/mL tetracycline. The medium is changed every 2 to 3 days and passed on for 1 to 2 generations for treatment. In recent years, it has been reported that 4-fluoro, 2-hydroxyquinoline (Ciprofloxacin, Cip), Pleu-romutilin derivative (Pleu-romutilin derivative, BM-Cyclin2: BM-1 and tetracycline derivative (BM-2)) Antibiotics are effective in killing mycoplasma when used alone or in combination. These three antibiotics are all prepared into 250X concentrated solutions in PBS and stored at -20°C for later use. The usage concentration Cip is 10 μg/mL, BM-1 is 10 μg/mL, and BM-2 is 5μg/mL. When using, first aspirate the contaminated culture medium, add RPMI1640 culture medium containing BM-1, then aspirate the culture medium after 3 days, add RPMI1640 culture medium containing BM-2, and culture for 4 days, and so on for 3 consecutive days. rounds, until it is proved by 33258 fluorescent staining microscopy that mycoplasma has been eliminated, then normal culture medium is added for culture and passage 3-4 times.
2. Heating treatment
Incubating contaminated tissue culture at 41°C for 18 hours can kill mycoplasma, but has adverse effects on cells. Therefore, a preliminary test should be conducted before treatment to explore the heating time that can kill mycoplasma to the maximum extent and have the least impact on cells. This method is sometimes unreliable. If treated with drugs first and then heated at 41°C, the effect will be better.

Cell Factory
3. Use mycoplasma-specific serum
Mycoplasma contamination can be removed with 5% rabbit mycoplasma immune serum (hemagglutination titer 1:320 or above). Because the specific antibody can inhibit the growth of mycoplasma, it turns negative 11 days after antiserum treatment and remains negative 5 months later. is negative. However, this method is more troublesome and not as convenient and economical as using antibiotics.
4. Other methods
In addition to the above-mentioned methods of removing contamination, there are also inoculation and sterilization methods in animals, macrophage phagocytosis methods, methods of adding bromouracil to contaminated culture bottles and then irradiating them with light, and filtration methods, etc., but they are all more troublesome and ineffective. Therefore, once mycoplasma contamination occurs, unless it is of particularly important value, it is generally discarded and re-cultured.
0 notes
Text
FG Troches atau FG Troches Meiji adalah antibiotik berbentuk tablet isap yang bermanfaat untuk mengatasi infeksi bakteri pada rongga mulut, seperti radang tenggorokan atau sariawan. Obat ini juga digunakan untuk mencegah terjadinya infeksi setelah prosedur operasi pada mulut.
FG Troches mengandung neomycin sulfate (fradiomycin sulfate) 2,5 mg dan gramicidin 1 mg. Kedua antibiotik ini bekerja menghentikan pertumbuhan dan penyebaran bakteri, seperti bakteri Streptococcus atau Bacteroides, yang dapat menyebabkan infeksi di rongga mulut dan tenggorokan. domino99
FG Troches tersedia dalam bentuk tablet isap. Obat ini dapat dikonsumsi oleh anak-anak dan orang dewasa. Namun, perlu diketahui bahwa gejala infeksi di rongga mulut, seperti radang tenggorokan, sariawan, atau gusi bengkak, umumnya disebabkan oleh virus dan dapat sembuh dengan sendirinya tanpa pengobatan.
Penggunaan antibiotik yang tidak tepat dapat menyebabkan resistensi antibiotik. Hal ini membuat infeksi bakteri jadi lebih sulit untuk diobati. Oleh karena itu, penggunaan antibiotik apa pun, termasuk FG Troches, perlu dikonsultasikan dengan dokter terlebih dahulu.
Apa Itu FG Troches
Golongan Obat resep
Kategori Antibiotik
Manfaat Mengobati sariawan, radang gusi, radang amandel, atau radang tenggorokan akibat infeksi bakteri
Dikonsumsi oleh Dewasa dan anak-anak
FG Troches untuk ibu hamil dan menyusui Kategori N: Belum diketahui.
Antibiotik dalam FG Troches bekerja secara lokal (di rongga mulut dan tenggorokan) dalam dosis yang rendah. Oleh karena itu, kecil kemungkinan obat ini terserap ke aliran darah dan menyebabkan efek samping pada janin.
Namun, perlu diketahui bahwa neomycin dosis normal yang diminum terbukti dapat menyebabkan efek samping pada janin. Jika Anda sedang hamil, sebaiknya konsultasikan mengenai penggunaan obat ini kepada dokter.
Karena hanya bekerja secara lokal, kandungan antibiotik dalam FG Troches kemungkinan tidak dapat terserap ke dalam ASI. Namun, tetap konsultasikan penggunaan obat ini dengan dokter jika Anda sedang menyusui.
Bentuk obat Tablet isap
Peringatan Sebelum Mengonsumsi FG Troches
FG Troches merupakan antibiotik yang tidak boleh dikonsumsi secara sembarangan. Berikut adalah hal yang perlu Anda perhatikan sebelum mengonsumsi FG Troches:
Jangan mengonsumsi FG Troches jika Anda alergi dengan kandungan dalam obat ini atau aminoglikosida lain, seperti amikacin, gentamicin, kanamycin, streptomycin, atau tobramycin.
Jangan mengonsumsi FG Troches lebih dari 7 hari, kecuali atas petunjuk dokter. Konsultasikan dengan dokter jika keluhan yang Anda alami belum membaik dengan FG Troches setelah 7 hari penggunaan.
Beri tahu dokter jika Anda menderita penyakit ginjal, myasthenia gravis, penyakit Parkinson, kolitis ulseratif, penyakit Crohn, atau irritable bowel syndrome.
Konsultasikan dengan dokter perihal penggunaan FG Troches jika Anda sedang mengonsumsi obat, suplemen, atau produk herbal tertentu, untuk mengantisipasi interaksi antarobat yang mungkin terjadi.
Konsultasikan dengan dokter perihal penggunaan FG Troches jika Anda sedang hamil atau menyusui.
Segera ke dokter jika mengalami reaksi alergi obat setelah mengonsumsi FG Troches.
Dosis dan Aturan Pakai FG Troches
Dosis FG Troches yang diresepkan oleh dokter tergantung pada kondisi pasien. Berikut adalah rincian dosis umum FG Troches:
Dewasa: 1–2 tablet isap, 4–5 kali sehari.
Anak-anak: 1 tablet isap, 4–5 kali sehari.
Cara Mengonsumsi FG Troches dengan Benar
Ikuti anjuran dokter dan baca informasi yang tertera pada kemasan FG Troches sebelum mulai mengonsumsinya. Jangan mengurangi atau menambah dosis tanpa arahan dari dokter.
FG Troches dapat dikonsumsi saat perut kosong atau sesudah makan. Isap tablet FG Troches di dalam mulut sampai habis, seperti mengisap permen. Pastikan untuk banyak minum air putih selama mengonsumsi FG Troches. Pastikan ada jarak waktu yang cukup antara satu dosis dengan dosis berikutnya.
FG Troches merupakan antibiotik. Pastikan Anda menghabiskan seluruh dosis FG Troches yang diresepkan oleh dokter meskipun kondisi Anda sudah membaik. Hal itu dilakukan untuk mencegah bakteri kebal terhadap antibiotik.
Simpan FG Troches dalam wadah tertutup di tempat yang kering dan sejuk. Hindarkan obat dari paparan sinar matahari dan jauhkan obat dari jangkauan anak-anak.
Interaksi FG Troches dengan Obat Lain
Belum diketahui secara pasti interaksi antarobat yang bisa terjadi jika FG Troches digunakan dengan obat lain. Melihat cara kerjanya, kecil kemungkinan obat ini dapat berinteraksi dengan obat lain. Interaksi obat baru mungkin terjadi jika FG Troches digunakan melebihi dosis yang dianjurkan.
Efek Samping dan Bahaya FG Troches
Efek samping yang berisiko muncul setelah mengonsumsi FG Troches yang mengandung neomycin adalah: domino77
Penurunan selera makan
Sakit perut
Mual
Muntah
Diare
Hubungi dokter jika efek samping yang diderita tidak kunjung membaik atau malah memburuk. Segera ke dokter jika terjadi reaksi alergi yang berat, seperti bengkak di bibir atau wajah, sesak napas, tenggorokan seperti tercekik, atau gatal-gatal di seluruh tubuh.
Lihat lebih lanjut mengenai:
Abses Peritonsil
Batu Amandel
Faringitis tangan judi rtp
0 notes
Text
OREGANO OIL AS PROMINENT NATURAL ANTIBIOTIC STUDY
Advertisement intended for healthcare professionals
Oxford Academic
Search Menu
Information
Account
Menu
Pathogens and Disease Federation of European Microbiological Societies
Article Navigation
Article Navigation
JOURNAL ARTICLE
Antibacterial effect of oregano essential oil alone and in combination with antibiotics against extended-spectrum β-lactamase-producing Escherichia coli
Hongbin Si, Jinqiang Hu, Zhichang Liu, Zhen-ling Zeng Author Notes
FEMS Immunology & Medical Microbiology, Volume 53, Issue 2, July 2008, Pages 190–194, https://doi.org/10.1111/j.1574-695X.2008.00414.x
Published: 01 July 2008 Article history
pdfPDF
Views
Cite
Permissions Icon Permissions
Share
Abstract
In this paper, we studied the antibacterial effects of oregano essential oil (OEO) both alone, using a twofold dilution method, and combined with antibiotics, using a checkerboard microtitre assay, against extended-spectrum β-lactamase (ESBL)-producing Escherichia coli. The result indicated that multiple drug-resistant E. coli was very sensitive to OEO and polymycin; their minimal inhibitory concentration values are 0.5 µL mL−1 and 0.8 µg mL−1. The antibacterial effects of OEO in combination with kanamycin were independent against E. coli, with fractional inhibitory concentration (FIC) indices of 1.5. The antibacterial effects of OEO combined with amoxicillin, polymycin, and lincomycin showed an additive effect against E. coli, with FIC indices in the range of 0.625–0.750. The antibacterial effects of OEO in combination with fluoroquinolones, doxycycline, lincomycin, and maquindox florfenicol displayed synergism against E. coli, with FIC indices ranging from 0.375 to 0.500. The combination of OEO with fluoroquinolones, doxycycline, lincomycin, and maquindox florfenicol to treat infections caused by ESBL-producing E. coli may lower, to a great extent, the effective dose of these antibiotics and thus minimize the side effects of antibiotics. This is the first report on OEO against ESBL-producing E. coli.
oregano essential oil, ESBLs, combination
Issue Section: research articles
Introduction
With the misuse and overuse of antibiotics to treat diseases, resistance to the drugs has begun to appear and has become more serious because of selective pressure. Resistance to multiple drugs is common for some kinds of clinically isolated bacterial strains; one of the mechanisms in multiple drug resistance is that bacteria can produce extended-spectrum β-lactamases (ESBLs) (Jacoby & Medeiros, 1991; Perilli et al., 1997). ESBL-producing Enterobacteriaceae and other kinds of bacteria have been reported widely. Infections caused by ESBL-producing bacteria have become a clinical and therapeutic problem because these organisms are resistant not only to β-lactamases but also to many other antimicrobial agents (Velasco et al., 2007). Often, ESBL-producing bacterial strains were isolated from animals in the veterinary field as well. It is important to prevent bacteria from producing ESBLs and to control diseases caused by ESBL-producing pathogens. The best way of treating these diseases is with imipenem and meropenem and third-generation cephalosporins in combination with enzyme inhibitors; however, these drugs cannot be used extensively in the veterinary field because of their high prices. Therefore, more economical and effective antibacterial agents are urgently needed in the veterinary field.
A feasible approach to limiting the transmission of these pathogens is to use essential oils as alternative agents or topical agents. Oregano, one kind of labiate Origanum plant that has been known for a long time as a popular remedy, is a very versatile plant. It was reported that Origanum compactum, Origanum minutif lorum, and Origanum majorana exhibit antifungal activity, antibacterial activity, and antimicrobial activity, respectively (Bouchra et al., 2003; Baydar et al., 2004; Vági et al., 2005). Until now its potential therapeutic roles such as diaphoretic, carminative, antispasmodic, antiseptic, and tonic properties have been recognized (Nostro et al., 2004). Sometimes OEO in conjunction with VP/MAP at chill temperatures can be used as a means of controlling the spoilage and safety of meat (Skandamis et al., 2002). In addition, it has been used widely in China as a kind of feed additive because it has a broad spectrum of action against bacteria, a rapid effect, and little residue, and now its antibacterial effect has been researched in vitro (Sivropoulou et al., 1996; Dorman & Deans, 2000; Force et al., 2000; Aligiannis et al., 2001; Lambert et al., 2001; Manohar et al., 2001) and in vivo (Adam et al., 1998) throughout the world. There is a lack of knowledge about OEO, which is one kind of volatile oil of Origanum vulgare linn (O. vulgare L.), used against ESBL-producing E. coli isolated from poultry in the veterinary field, especially on its combination with other antibacterial agents. The mechanism may be in its principal components, i.e. thymol and carvacrol, which have antimicrobial properties.
The objective of the present work was to investigate the antibacterial effects of OEO against ESBL-producing E. coli. The effects of OEO in combination with antibiotics were also evaluated in order to seek effective combinations to treat emergent infections caused by ESBL-producing E. coli. This is the first report on OEO alone and in combination with antibacterial agents against ESBL-producing E. coli isolated from chicken.
Materials and methods
Bacterial strains
Escherichia coli isolates were obtained from chicken livers from a chicken farm at Henan Agricultural University in Zhengzhou, Henan Province, China. Strains were isolated and purified on an agar plate and identified using a Vitek 32 system and GNI+card (bioMérieux, France).
Screening for and confirmation of ESBLs
To detect the ESBLs, the initial screening was performed by testing the zone diameter for ceftriaxone (30 µg), ceftazidime (30 µg), cefotaxim (30 µg), and aztreonam (30 µg) and interpreted by the criterion of NCCLS (Adam et al., 1998; Aligiannis et al., 2001), where a positive result was considered to be suspicious for the presence of ESBLs. This test was followed by a phenotypic confirmation test of ESBLs, carried out by testing the zone diameter for cefotaxim (30 µg) and ceftazidime (30 µg) with and without clavulanate (10 µg), as described previously by the NCCLS. A 5 mm increase in the zone diameter for ceftazidime or cefotaxim tested in combination with clavulanate vs. the zone when tested alone was considered indicative of ESBL production. Klebsiella pneumoniae American Type Culture Collection (ATCC) 700603 and E. coli ATCC 25922 were used as positive and negative controls, respectively (Adam et al., 1998; Dorman & Deans, 2000).
Identification of ESBLs gene
Templates of total DNA from the isolate were prepared as described previously and the identity of the ESBLs was determined by PCR amplification of selected determinants followed by direct sequencing of the amplicons as described previously. The primers used for detecting ESBLs−TEM genes were as follows: the forward primer (5′-GGGGATGAGTATTCAACATTTCC-3′) and the reverse primer (5′-GGGCAGTTACCAATGCTTAATCA-3′) were designed using OLIGO 6.0 software based on the published ESBL gene sequence (Genbank accession no. AF332513). The sequence region of an 861-bp fragment containing an ORF of a full-length ESBL gene was predicted between the two primers. PCR was carried out in a final volume of 50 µL containing 1 µg template DNA, 100 pmol of each primer (flo1/flo2), 1 × PCR buffer, 0.2 mM of each of the dNTPs, and 2.5 U Ex Taq polymerase (Takara, Japan). A total of 30 cycles were performed in the PCR Express (HYBRID Corporation), under the following conditions: denaturation at 94 °C for 1 min, annealing at 53 °C for 30 s, and extension at 72 °C for 1 min with final termination at 72 °C for 10 min.
The PCR products were electrophoresed on 1.0% agarose gel. The DNA band of interest was excised, purified, and ligated to pGEM-T Easy vector (Promega), and the recombinant vector named pGEM-ESBL was constructed. JM109 competent cells were transformed, and the white clones were screened and digested with restriction enzyme EcoRI and sequenced for identification.
Oregano essential oil (OEO)
OEO was collected as previously described (Nostro et al., 2004). The aerial parts of oregano (O. vulgare L.), obtained from a commercial source (Yuanzheng Ltd, Hebei, China), were subjected to hydrodistillation for 2 h using a modified clevenger type apparatus, and the essential oil was collected and stored at 4 °C. All oils were diluted (v/v) by both the agar and broth dilution methods.
Determination of minimal inhibitory concentration (MIC)
The isolated strain was stored at −70 °C until use; E. coli was grown in Martin broth modified and adjusted with medium to match the 0.5 McFarland standard (105 CFU mL−1). The MICs of OEO, defined as the lowest concentration that completely inhibited visible bacterial growth after 14 h, and antibiotics were examined by broth dilution method in 96-well, microtitre plates. The MICs of the agents sarafloxacin, levofloxacin, polymycin, lincomycin, amoxicillin, ceftiofur, ceftriaxone, maquindox, florfenicol, doxycycline, and kanamycin against the isolated strain were determined by the twofold dilution method, and the results were interpreted in accordance with the recommendations of the National Committee for Clinical Laboratory standards (Adam et al., 1998; Dorman & Deans, 2000). Escherichia coli strain ATCC 25922 was used as quality control in MIC determination.
Organisms were measured by twofold dilutions of OEO samples with Martin broth modified solutions containing 0.5% Tween-80 to enhance oil solubility. The concentration of essential oil in the medium ranged from 0.5% to 0.0039% (v/v). All determinations were performed in triplicate. If MIC values were different, the trial was repeated until the values were the same. In this way, the result is certain. Two growth controls consisting of Martin broth modified medium and Martin broth modified with 1.0% Tween-80 were added to ascertain that these vehicles did not affect bacterial growth.
Checkerboard microtitre test
Ten serial, twofold dilutions of the bacterial fraction and essential oil were prepared using the same solvents as in the MIC tests. Aliquots (50 µL) of antibacterial dilution were added to the wells of a 96-well plate in a vertical orientation and 10 µL aliquots of OEO dilution were added in a horizontal orientation so that the plate could contain various concentration combinations of the two compounds. Then, each well was inoculated with 100 µL (c. 5 × 105 CFU well−1) of antibacterial bacterial suspensions and cultivated at 37 °C. Fractional inhibitory concentration (FIC) was calculated by dividing the MIC of the combination of antibacterial and essential oil by the MIC of antibacterial or essential oil alone. The FIC index, obtained by adding both FICs, was interpreted as synergistic when it was ≤0.5, as additive (indifferent) when it was >0.5 and ≤2.0, and as antagonistic when it was >2.0 (White et al., 1996). Similar checkerboard experiments were performed as well to test the combined effect of the essential oil with each antibacterial.
Results and discussion
The initial screening was carried out to test inhibition zones of ceftriaxone, ceftazidime, cefotaxim and aztreonam, and the corresponding inhibitory diameters were 14 mm, 16 mm, 17 mm, and 14 mm, respectively, which were consistent with the criterion of NCCLS. Thus, it was tentative that this strain might produce >ESBLs. Moreover, the confirmation test of ESBL production indicated that the inhibition zones of cefotaxim and cefotaxime-plus-clavulanate disks, ceftazidime and ceftazidime-plus-clavulanate disks were 14 mm, 20 mm, 15 mm and 17 mm, respectively. A distance of 6 mm between the zones of cefotaxim and cefotaxim-plus-clavulanate disks was considered to indicate the production of ESBLs. Besides, the resulting PCR product of the ESBLs−TEM gene was about 861 bp in length and identical to the expected size. Further identification of the restriction enzyme and sequencing of recombinant plasmid pGEM-ESBLs undoubtedly confirmed that the isolate carried the ESBLs-TEM gene. Sequence analysis of the 861-bp amplicon produced with the TEM-specific primers showed that the plasmid harbored a TEM-type gene with 100% identity to TEM-116 (AJ847364); the result revealed that the 287-amino-acid protein was the same type gene with 100% identity to TEM-116 (AJ847364). TEM-116 is one of primary, prevalent ESBL types. It is remarkable that there is 22.1% TEM-116 in a survey of ESBL-producing pathogens in a Spanish University Hospital; especially 49.32% TEM-116 pathogens can produce other ESBL types at the same time (Romero et al., 2007). More than 400 different types of β-lactamases originating from clinical isolates have been described. Since they were first identified at the beginning of the 1980s (Perilli et al., 1997), ESBL-producing Enterobacteriaceae have been reported widely (Jarlier et al., 1988; Sirot et al., 1988; Jacoby & Medeiros, 1991; Yang et al., 1998; Bonnet et al., 1999). ESBL occurred predominantly not only in Klebsiella species and E. coli but also in other genera of the family Enterobacteriaceae, such as Shigella and others (Ashkenazi et al., 2003; Ahmed et al., 2004). Organisms producing ESBLs are clinically relevant and remain an important cause for failure of therapy with cephalosporins. It is very important to find some effective drugs against these diseases.
The susceptibility test results of the drug alone suggested that ESBLs-producing E. coli had serious resistance to commonly used antibacterial agents including fluoroquinolones, doxycycline, lincomycin, maquindox, florfenicol, and amoxicillin, and most of their MIC values were over 16 µg mL−1, indicating that the ESBL-producing E. coli had serious resistance to multiple drugs. This result was consistent with some recent surveys that ESBL-producing E. coli exhibited cross-resistance to tetracycline, gentamicin, and ciprofloxacin. ESBL-producing pathogens are often encoded by genes located on large plasmids, and these also carry a gene for resistance to other antibiotics (Bradford, 2001; Rahal, 2002).
Thus, this would probably bring many difficulties in treating diseases caused by these bacteria. However, this strain was sensitive to polymycin with the MIC value of 0.8 µg mL−1, implying that polymycin could treat the infection disease caused by these drug-resistant pathogens. It should be considered that polymycin was used to treat these diseases alone because of its poor oral absorption. Viewed from the susceptibility test result, ESBL-producing bacteria were sensitive to OEO. Moreover, the bacteria were still sensitive to the same strain when the original antibacterial compound was diluted 10 000-fold, suggesting that it was worthwhile to explore an effective alternative to antibiotics to treat diseases caused by bacteria, especially ESBL-producing bacteria.
The efficacy of OEO combined with an antibacterial agent was determined by checkboard assay, and the results are listed in Table 1. The MIC value of OEO combined with many antibacterial agents decreased predominantly, suggesting that OEO was synergistic. This is the first report concerning the synergistic effects of OEO in combination with antibiotics against bacteria, especially ESBL-producing bacteria. The MIC value of OEO in combination with most of the bacterial agents against ESBL-producing E. coli remarkably decreased, with FIC indices ranging from 0.375 to 0.75 (excluding the combination with kanamycin with an FIC index of 1.5. The FIC index indicated an independence between oil and kanamycin against ESBL-producing E. coli.) Moreover, the susceptibilities of bacteria to amoxicillin, polymycin, lincomycin, and third-generation cephalosporin were enormously improved by combination with OEO. The FIC indices of OEO combined with polymycin, lincomycin, amoxicillin, and cephalosporins were 0.75, 0.75, 0.75, and 0.625, respectively, and these data showed that these combinations had an additive effect. FIC indices of OEO combined with fluoroquinolones, doxycycline, lincomycin, maquindox, and florfenicol altered between 0.375 and 0.5. Hence the FIC indices of OEO combined with these antibiotics were synergistic.
Table 1Concentrations of OEO (µL mL−1) and of antibacterial (µg mL−1)
Drugs Single drug Combined use of drugs FICs Results
OEO 0.5 0.125 0.375 Synergism
Sarafloxacin 640 80
OEO 0.5 0.125 0.5 Synergism
Levofloxacin 64 16
OEO 0.5 0.125 0.75 Additive effect
Polymycin 1.6 0.8
OEO 0.5 0.125 0.75 Additive effect
Lincomycin 320 160
OEO 0.5 0.25 0.75 Additive effect
Amoxicillin 1280 320
OEO 0.5 0.25 0.625 Additive effect
Ceftiofur 640 80
OEO 0.5 0.25 0.625 Additive effect
Ceftriaxone 640 80
OEO 0.5 0.125 0.5 Synergism
Maquindox 64 16
OEO 0.5 0.125 0.375 Synergism
Florfenicol 320 40
OEO 0.5 0.125 0.375 Synergism
Doxycycline 64 16
OEO 0.5 0.25 1.5 Independence
Kanamycin 1280 1280
FIC, fractional inhibitory concentration; OEO, oregano essential oil.
Open in new tab
Although an effect of oregano on chicken lactobacilli and E. coli was studied recently (Horosová, 2006), little was known about the mechanism of action for OEO on different bacteria or about the specific effects of OEO constituents, e.g. carvacrol and thymol (Cox et al., 1998; Helander et al., 1998; Ultee et al., 1999; Skandamis et al., 2000; Tassou et al., 2000). According to Conner & Beuchat (1984), the antimicrobial action of essential oils might be due to the impairment of a variety of enzyme systems, including those involved in energy production and structural component synthesis. Overall, we can conclude that OEO is one of the most promising natural compounds that can be used to develop safer antibacterial agents and that its effective combination with antibacterials may be used in the future to treat diseases caused by ESBL-producing E. coli in the veterinary field instead of expensive antibacterial agents.
Acknowledgement
This work was supported by The Research Fund for the Doctoral Program of Higher Educator (RFDP, No. 30471307).
References
Adam K. Sivropoulou A. Kokkini S. Lanaras T. Arsenakis M. (1998) Antifungal activity of Origanum vulgare subsp. hirtum, Mentha spicata, Lavandula angustifolia, and Salvia fruticosa essential oils against human pathogenic fungi. J Agric Food Chem46: 1739–1745.
Google ScholarCrossrefWorldCat
Ahmed A.M. Nakano H. Shimamoto T. (2004) The first characterization of extended-spectrum beta-lactamase-producing Salmonella in Japan. J Antimicrob Chemother54: 283–284.
Google ScholarCrossrefPubMedWorldCat
Aligiannis N. Kalpoutzakis E. Mitaku S. Chinou B.I. (2001) Composition and antimicrobial activity of the essential oils of two Origanum species. J Agric Food Chem49: 4168–4170.
Google ScholarCrossrefPubMedWorldCat
Ashkenazi S. Levy I. Kazaronovski V. Samra Z. (2003) Growing antimicrobial resistance of Shigella isolates. J Antimicrob Chemother51: 427–429.
Google ScholarCrossrefPubMedWorldCat
Baydar H. Sagdic OZkan G. et al. . (2004) Antibacterial activity and composition of essential oils from Origanum, Thymbra and Satureja species with commercial importance in Turkey. Food Control15: 169–172.
Google ScholarCrossrefWorldCat
Bonnet R. De Champs C. Sirot D. Labia R. Sirot J. (1999) Diversity of TEM mutants in Proteus mirabilis. Antimicrob Agents Ch43: 2671–2677.
Google ScholarWorldCat
Bouchra C. Achouri M. Idrissi Hassani L.M. et al. . (2003) Chemical composition and antifungal activity of essential oils of seven Moroccan Labiatae against Botry tiscinerea Pers:Fr. J Ethnopharmacol89: 165–169.
Google ScholarCrossrefPubMedWorldCat
Bradford P.A. (2001) Extended-spectrum β-lactamases in the 21st century: characterization, epidemiology, and detection of this important resistance threat. Clin Microbiol Rev14: 933–951.
Google ScholarCrossrefPubMedWorldCat
Conner D.E. Beuchat L.R. Worthington R.E. Hitchcock H.L. (1984) Effects of essential oils and oleoresins of plants on ethanol production, respiration and sporulation of yeasts. Int J Food Microbiol1: 63–74.
Google ScholarCrossrefWorldCat
Cox S.D. Gustafson J.E. Mann C.M. Markham J.L. Liew Y.C. Hartland R.P. Bell H.C. Warmington J.R. Wyllie S.G. (1998) Tea tree oil causes K+ leakage and inhibits respiration in Escherichia coli. Lett Appl Microbiol26: 355–358.
Google ScholarCrossrefPubMedWorldCat
Dorman HJD Deans S.G. (2000) Antimicrobial agents from plants: antibacterial activity of plant volatile oils. J Appl Microbiol88: 308–316.
Google ScholarCrossrefPubMedWorldCat
Force M. Sparks W.S. Ronzio A. (2000) Inhibition of enteric parasites by emulsified oil of oregano in vivo. Phytother Res14: 213–214.
Google ScholarCrossrefPubMedWorldCat
Helander I.K. Alakomi H.L. Latva-Kala K. Mattila-Sandholm T. Pol I. Smid E.J. Von Wright A. (1998) Characterization of the action of selected essential oil components on Gram negative bacteria. J Agric Chem46: 3590–3595.
Google ScholarCrossrefWorldCat
Horosová K. Bujnáková D. Kmet V. (2006) Effect of oregano essential oil on chicken lactobacilli and E. coli. Folia Microbiol (Praha)51: 278–280.
Google ScholarCrossrefPubMedWorldCat
Jacoby G.A. Medeiros A.A. (1991) More extended-spectrum β-lactamases. Antimicrob Agents Chemother35: 1697–1704.
Google ScholarCrossrefPubMedWorldCat
Jarlier V. Nicolas M.H. Fournier G. Philippon A. (1988) Extended broad-spectrum β-lactamases conferring transferable resistance to newer β-lactam agents in Enterobacteriaceae: hospital prevalence and susceptibility patterns. Rev Infect Dis10: 867–878.
Google ScholarCrossrefPubMedWorldCat
Lambert RJW Skandamis P.N. Coote P.J. Nychas GJE (2001) A study of the minimum inhibitory concentration and mode of action of oregano essential oil, thymol and carvacrol. J Appl Microbiol91: 453–462.
Google ScholarCrossrefPubMedWorldCat
Manohar V. Ingram C. Gray J. Talpur N.A. Echard B.W. Bagchi D. Preuss G. (2001) Antifungal activities of origanum oil against Candida albicans. Mol Cell Biochem228: 111–117.
Google ScholarCrossrefPubMedWorldCat
Nostro A. Blanco A.R. Cannatelli M.A. Enea V. Flamini G. Morelli I. Roccaro A.S. Alonzo V. (2004) Susceptibility of methicillin-resistant staphylococci to oregano essential oil, carvacrol and thymol. FEMS Microbiol Lett230: 191–195.
Google ScholarCrossrefPubMedWorldCat
Perilli M. Felici A. Franceschini N. De Santis A. Pagani L. Luzzaro F. Oratore A. Rossolini G.M. Knox J.R. Amicosante G. (1997) Characterization of a new TEM-derived β-lactamase produced in a Serratia marcescens strain. Antimicrob Agents Chemother41: 2374–2382.
Google ScholarPubMedWorldCat
Rahal J.J. Urban C. Segal-Maurer S. (2002) Nosocomial antibiotic resistance in multiple gram-negative species: experience at one hospital with squeezing the resistance balloon at multiple sites. Clin Infect Dis34: 499–503.
Google ScholarCrossrefPubMedWorldCat
Romero EDV Padilla T.P. Hernández A.H. Grande R.P. Vázquez M.F. García I.G. García-Rodríguez J.A. Muñoz Bellido J.L. (2007) Prevalence of clinical isolates of Escherichia coli and Klebsiella spp. producing multiple extended-spectrum β-lactamases. Diagn Microbiol Infect Dis59: 433–437.
Google ScholarCrossrefPubMedWorldCat
Sirot J. Chanal C. Petit A. Sirot D. Labia R. Gerbaud G. (1988) Klebsiella pneumoniae and other Enterobacteriaceae producing novel plasmid-mediated β-lactamases markedly active against third-generation cephalosporins: epidemiologic studies. Rev Infect Dis10: 850–859.
Google ScholarCrossrefPubMedWorldCat
Sivropoulou A. Papanicolau E. Nicolaou C. Kokkini S. Lanaras T. Arsenakis M. (1996) Antimicrobial and cytotoxic activities of Origanum essential oils. J Agric Food Chem44: 1202–1205.
Google ScholarCrossrefWorldCat
Skandamis P. Tsigarida E. Nychas GJE (2000) Ecophysiological attributes of Salmonella typhimurium in liquid culture and within gelatin gel with or without the addition of oregano essential oil. World J Microbiol Biotechnol16: 31–35.
Google ScholarCrossrefWorldCat
Skandamis P. Tsigarida E. Nychas G-JE (2002) The effect of oregano essential oil on survival/death of Salmonella typhimurium in meat stored at 5 °C under aerobic, VP/MAP conditions. Food Microbiol19: 97–103.
Google ScholarCrossrefWorldCat
Tassou C.C. Koutsoumanis K. Nychas GJE (2000) Inhibition of Salmonella enteritidis and Staphylococccus aureus in nutrient broth by mint essential oil. Food Res Intern33: 273–280.
Google ScholarCrossrefWorldCat
Ultee A. Kets EPW Smid E.J. (1999) Mechanisms of action of carvacrol on the food-borne pathogen Bacillus cereus. Appl Environ Microbiol65: 4606–4610.
Google ScholarPubMedWorldCat
Vági E. Simándi B. Suhajda Á Héthelyi É (2005) Essential oil composition and antimicrobial activity of Origanum majorana L. extracts obtained with ethyl alcohol and supercritical carbon dioxide. Food Res Int38: 51–57.
Google ScholarCrossrefWorldCat
Velasco C. Romero L. Martínez JMR Rodríguez-Baño J. Pascual A. (2007) Analysis of plasmids encoding extended-spectrum β-lactamases (ESBLs) from Escherichia coli isolated from non-hospitalised patients in Seville. Int J Antimicrob Agents29: 89–92.
Google ScholarCrossrefPubMedWorldCat
White R.L. Burgess D.S. Manduru M. Bosso J.A. (1996) Comparison of three different in vitro methods of detecting synergy: time-kill, checkerboard, and E test. Antimicrob Agents Chemother40: 1914–1918.
Google ScholarPubMedWorldCat
Yang Y. Bhachech N. Bradford P.A. Jett B.D. Sahm D.F. Bush K. (1998) Ceftazidime-resistant Klebsiella pneumoniae and Escherichia coli isolates producing TEM-10 and TEM-43 β-lactamases from St. Louis, Missouri. Antimicrob Agents Chemother42: 1671–1676.
Google ScholarPubMedWorldCat
Author notes
Editor: Willem van Eden
© 2008 Federation of European Microbiological Societies
CITATIONS
75 75 total citations on Dimensions.
VIEWS
43,621
ALTMETRIC
Article has an altmetric score of 7
Information More metrics information
Email alerts
Article activity alert
Advance article alerts
New issue alert
In progress issue alert
Receive exclusive offers and updates from Oxford Academic
Recommended
In vitro activity of temocillin against extended spectrum β-lactamase-producing Escherichia coli
Hector Rodriguez-Villalobos et al., Journal of Antimicrobial Chemotherapy, 2006
Evaluation of Oxoid combination discs for detection of extended-spectrum β-lactamases
De Gheldre et al., Journal of Antimicrobial Chemotherapy, 2003
Surveillance of antimicrobial resistance among Gram-negative isolates from intensive care units in eight hospitals in Turkey
Study group Sebahat Aksaray et al., Journal of Antimicrobial Chemotherapy, 2000
A silent network’s resounding success: how mutations of core metabolic genes confer antibiotic resistance
Gamal Wareth et al., Selections from Signal Transduction and Targeted Therapy, 2021
A review on catalytic & non-catalytic bio-oil upgrading in supercritical fluids
Sainab Omar et al., Frontiers of Chemical Science and Engineering, 2021
Rock Types and Reservoir Characteristics of Shahejie Formation Marl in Shulu Sag, Jizhong Depression, Bohai Bay Basin
Jingwei Cui et al., Journal of Earth Science, 2021
Powered by
Citing articles via
Web of Science (66)
Google Scholar
Latest
Most Read
Most Cited
Comparative study of GBP recruitment on two cytosol-dwelling pathogens, Francisella novicida and Shigella flexneri highlights differences in GBP repertoire and in GBP1 motif requirements
Murine herpesvirus-68-related growth factors treatment correlates with decrease of p53 and HIF-1α protein levels
The Burkholderia contaminans prevalent phenotypes as possible markers of poor clinical outcomes in chronic lung infection of children with cystic fibrosis
Induction and sustenance of antibacterial activities distinguishes response of mice to Salmonella Typhi from response to Salmonella Typhimurium
In vitro and in vivo evaluation of DNase I in reinstating antibiotic efficacy against Klebsiella pneumoniae biofilms
More from Oxford Academic
Biological Sciences Microbiology Science and Mathematics Books Journals
About Pathogens and Disease
Editorial Board
Policies
Author Guidelines
Contact Us
Facebook
Twitter
LinkedIn
Purchase
Recommend to your Library
Advertising and Corporate Services
Journals Career Network
Pathogens and Disease
Online ISSN 2049-632X
Copyright © 2023 Federation of European Microbiological Societies
About Oxford Academic
Publish journals with us
University press partners
What we publish
New features
Authoring
Open access
Purchasing
Institutional account management
Rights and permissions
Get help with access
Accessibility
Contact us
Advertising
Media enquiries
Oxford University Press
News
Oxford Languages
University of Oxford
Oxford University Press is a department of the University of Oxford. It furthers the University's objective of excellence in research, scholarship, and education by publishing worldwide
Oxford University Press
Copyright © 2023 Oxford University Press Cookie policy Privacy policy Legal notice
0 notes
Photo

Multidrug Resistant Acinetobacter baumannii. This Gram-negative bacteria came home with Iraq and Afghanistan veterans and has become a source of hospital-acquired infection. Treat with sulbactam, aminoglycosides, polymixyns, or tigecycline. **Cystic fibrosis and lung transplants**
MDR = Multi-Drug-Resistant Tuberculosis. When TB is resistant to the two most potent anti-TB drugs, isoniazid and rifampin (and maybe others). Treat with amikacin, kanamycin, or capreomycin for up to two years.
XDR = Extensively Drug-Resistant Tuberculosis. A rare form (10%) of MDR TB resistant to isoniazid and rifampin, all fluoroquinolones, and at least one of three injectable second-line drugs (amikacin, kanamycin, or capreomycin). Requires at least 6 drugs for acute treatment and 4 for maintenance.
1 note
·
View note
Last Seen Blogs

fez-pwned
fez 🦭

leslilli
My Green Cauldron

alternate5
Untitled

aprilseojinlee
April Seojin Lee

s-and-n
s and n